Дефицит природных киллеров: гетерогенность, клиника, диагностика, лечение, клинические приложения
Резюме.
Дефицит природных киллеров - это обобщающий термин, который применяется для обозначения группы гетерогенных по происхождению болезней иммунной системы, при которых отмечается уменьшенное количество и/или нарушенная функциональная активность субпопуляции лимфоцитов с фенотипом CD3-CD16+CD56+, однако имеют место нормальные другие показатели иммунной системы. Можно выделить первоначальный и вторичный дефицит естественных киллеров. В настоящее время описано более 10 первичных форм этого иммунодефицита. В клинике преобладают вирусные инфекции, аутоиммунные и неопластические осложнения, а также некоторые дополнительные симптомы. Для диагностики употребляют проточную цитометрию, цитологический анализ и ряд функциональных тестов. В качестве базовой терапии применяют препараты интерферонов и рекомбинантного интерлейкина-2. Приведены два клинических случая из собственной практики. У одной пациентки с изолированным дефицитом природных киллеров развилась васкулопатия, вызванная вирусом ветряной оспы/опоясывающего герпеса, и лейкоэнцефалит, ассоциированный с вирусом герпеса человека 6-го типа. У другой больной сочетался дефицит С3-компонента комплемента и природных киллеров, что привело к хронической микст-инфекции, вызванной бета-гемолитическим стрептококком и вирусом Эпштейна-Барр, с соответствующими аутоиммунными осложнениями.
Вступление
Дефицит природных киллеров или природных киллерных клеток (natural killer (NK) cells) — это обобщающий термин, который применяется для обозначения группы гетерогенных по происхождению болезней иммунной системы, при которых отмечается уменьшенное количество и/или нарушенная функциональная активность субпопуляции лимфоцитов с фенотипом СD16+CD56+, однако имеют место другие нормальные показатели иммунного статуса (Orange J.S., 2006). Такие заболевания могут проявляться только симптомами иммунной недостаточности или сопровождаться некоторыми дополнительными проявлениями, не связанными с иммунной дисфункцией, что обусловлено плейотропными свойствами поврежденного гена. Можно выделить первичный и вторичный дефицит природных киллеров и различать количественные и качественные формы иммунодефицита. Генетически детерминированные варианты болезни отмечают чаще, поскольку факторы вторичной иммуносупрессии, такие как ионизирующая радиация, лучевая терапия, цитостатические химиопрепараты обычно оказывают более широкое влияние на иммунную систему, не ограничиваясь точечным влиянием на минорную субпопуляцию лимфоцитов.
Первичный дефицит природных киллеров относится к малым иммунодефицитным болезням, которым присуща большая распространенность в популяции, дебют в любом возрасте, длительные периоды бессимптомного течения, вариабельность клинической картины в течение жизни и гетерогенность симптомов даже в пределах одной семьи. Семейный дефицит природных киллеров впервые описан J.K. Portaro и соавторами (1978) как наследственный дефицит К-клеток у здоровых лиц. В дальнейшем выяснили, что с этим иммунодефицитом могут быть связаны клинические симптомы, иногда даже угрожающие жизни. Проявления болезни напрямую определяются функциональным назначением природных киллеров в организме человека. Эти клетки обусловливают противовирусное и противоопухолевое влияние благодаря участию в реакциях спонтанной и антителозависимой клеточно-опосредованной цитотоксичности, а также оказывают иммунорегуляторные воздействия путем продукции цитокинов, в частности интерферонов (ИФН). Поэтому в клинической картине преобладают частые, длительные, атипичные, аномально тяжелые или хронические вирусные инфекции, вызванные в основном агентами с оппортунистическими свойствами, а также злокачественные опухоли, преимущественно лимфопролиферативного происхождения. В то же время, природные киллеры не играют существенной роли во время острой инфекции, вызванной Toxoplasma gondii (Hughes H.P. et al., 1988). В последнее время изучаются адаптивные свойства природных киллеров, в частности способность развивать качественно более эффективный ответ при повторном контакте с антигеном (Sun J.C. et al., 2009), что сближает эти клетки с цитотоксическими Т-лимфоцитами.
Дефицит природных киллеров относится к потенциально курабельным болезням иммунной системы, поскольку разработано несколько видов иммунотерапевтических вмешательств, которые продемонстрировали свою эффективность в небольших контролируемых исследованиях. Это еще один из аргументов для активного выявления такой иммунной дисфункции.
Несмотря на большое количество сообщений в англоязычной литературе по этой проблеме, пока нет основательных обзоров, которые предоставляли бы исчерпывающую информацию о клинике, диагностике и лечении дефицита естественных киллеров человека. Более того, нами не выявлено ни одной отечественной статьи об этом иммунодефиците или случаях диагностики этой распространенной патологии иммунной системы человека в других клинических центрах Украины. Это значит, что люди с дефицитом природных киллеров не получают должного лечения, с чем может быть связано ухудшение качества и сокращение их продолжительности.
Эпидемиология
Пока нет точных статистических данных по распространению изолированного дефицита природных киллеров в человеческой популяции.
F. Nagao и соавторы (1995) провели популяционное исследование, изучив удельный вес дефицита
природных киллеров в двух группах наблюдения: среди здоровых доноров крови (n=68) и 57 больных автоиммунной патологией (n=36) и опухоли (n=21). Продемонстрировано, что количественный и функциональный дефицит клеток с фенотипом CD3-CD16+CD56+ отмечался в 23,3% случаев среди здоровых людей на момент обследования и в 70% - в группе больных. Терапия с помощью интерлейкина (ИЛ)-2 приводила к более быстрому восстановлению функции NK-клеток здоровых лиц с латентным иммунодефицитом, чем у иммуноскомпрометированных пациентов с клинической манифестацией.
G. Kabalak и соавторы (2010) в группе из 149 больных с синдромом Шегрена обнаружили гомозиготную делецию в гене иммуноглобулинообразного транскрипта 6-го типа (ILT6), кодирующего поверхностный рецептор естественных киллеров в 8% случаев, тогда как в группе здоровых людей =749) таковая мутация случалась только в 3% случаев.
I. Hwang и соавторы (2012) сообщили о дефиците сигнального адаптера FcR? при сохранении экспрессии CD3? в естественных киллерах трети обследованных добровольцев. Такое нарушение приводило к слабой экспрессии рецепторов NKp46 и NKp30, однако сохраненной экспрессии молекулы CD16. Подобные природные киллеры не демонстрировали реакции спонтанной цитотоксичности в отношении опухолевых клеток, однако даже активнее здоровых клеток принимали участие в антителозависимой цитотоксичности.
T. Ovesen и соавторы (2011) выявили дефицит природных киллеров у большинства из 18 пациентов с хронической посттимпанической отореей.
P. Bourin и соавторы (1993) изучили циркаденные ритмы, связанные с природными киллерами. Авторы выяснили, что каждая клетка осуществляет патрулирование организма примерно один раз в 24 часа, причем больше природных киллеров поступают в циркуляцию утром, а меньше всего - в ночное время.
Первичный дефицит NK-клеток встречается с одинаковой частотой у мужчин и женщин (Orange J.S., 2012), хотя прием пероральных контрацептивов может обусловить повышение частоты вторичных форм иммунодефицита у лиц слабого пола (Baker D.A. et al., 1989).
J. Kaplan и соавторы (1982) показали, что активность периферических природных киллеров у новорожденных ниже, чем у взрослых людей, однако под влиянием альфа-ИФН она быстро растет до уровня, даже превышающего таковой у взрослых людей.
D. Rukavina соавторы (1998) продемонстрировали снижение содержания перфорина в гранулах природных киллеров и цитотоксических Т-лимфоцитов у лиц старше 70 лет, особенно у мужчин. ИЛ-2 восстанавливал содержание этого белка в указанных клетках in vitro.
S. Vaquer и соавторы (1987) выявили пониженную функциональную активность природных киллеров у беременных и продемонстрировали эффективность ИЛ-2 и гамма-ИФН для потенциации функции NK-клеток in vitro в таких случаях.
Гетерогенность
N.E. Kay, J.M. Zarling (1984) описали особую форму дефицита природных киллеров у пациента, у которого впоследствии развился хронический лимфолейкоз. Эти клетки были лишены азурофильных гранул, в связи с чем не имели фенотипа больших гранулярных лимфоцитов, однако нормально экспрессировали поверхностные маркерные молекулы. Как известно, именно в азурофильных гранулах содержатся основные факторы агрессии природных киллеров – перфорин и гранзимы.
Мы также описали случай семейного дефицита крупных гранулярных лимфоцитов (Мальцев Д.В., 2009).
A. Komiyama и соавторы (1990) описали функциональный дефицит природных киллеров у сиблингов, который заключался в невозможности реализации реакций спонтанной клеточно-опосредованной цитотоксичности при сохранении антителозависимой цитотоксичности и функций Т-киллеров. Преобладали рецидивирующие респираторные инфекции. H. Matsuzaki и соавторы (1983) сообщили о противоположной ситуации, когда были нарушены реакции антителозависимой цитотоксичности, однако нормально реализовывалась спонтанная цитотоксичность, что видоизменяло клиническую картину, обусловливая развитие преимущественно аутоиммунной патологии.
E. de Vries и соавторы (1996) обнаружили мутацию в гене Fc? RIIIA, кодирующем молекулу CD16 природных киллеров. Имела место субституция T-to-A в позиции 230 в обеих аллелях, которая приводила к замене лейцина на гистидин в 48-й позиции первого экстрацеллюлярного иммуноглобулинообразного домена Fc? RIIIа. Мальчик страдал рецидивирующими респираторными вирусными инфекциями с рождения. У него отмечалось осложнение после вакцинации БЦЖ. Первичная инфекция, вызванная вирусом Эпштейна - Барр (Epstein-Barr virus - EBV), и ветряная оспа имели тяжелое течение. Часто рецидивировала инфекция, вызванная вирусом простого герпеса (herpes simplex virus HSV) 1-го типа (HSV-1). S. Jawahar и соавторы (1996) описали другую мутацию в том же гене — субституцию T-to-A в 230 позиции, что приводило к замене лейцина на гистидин в 66-й позиции первого иммуноглобулиноподобного домена молекулы CD16 в опознавательном сайте В73.1. У пациента отмечались частые респираторные вирусные инфекции и рецидивирующий орофациальный герпес.
A. Aoukaty и соавторы (2003) описали тяжелую хроническую активную EBV-инфекцию у мальчика с дефицитом LAIR-1, или CD305, относящегося к киллингингибирующим рецепторам природных киллеров. NK-клеток демонстрировали слабую цитотоксичность и способность к продукции гамма-ИФН.
R. Gazit и соавторы (2004) обнаружили измененную экспрессию KIR2DL1 природных киллеров, сопровождавшееся абсолютным ингибированием функции этих клеток у пациента, страдавшего рецидивирующими инфекциями, обусловленными вирусом ветряной оспы/опоясывающего герпеса (varicella-zoster virus) – CMV).
K.J. Maher и соавторы (2005) сообщили об изолированном дефиците перфорина в гранулах природных киллеров у некоторых пациентов с синдромом хронической усталости.
C. Eidenschenk и соавторы (2006a) описали новую форму изолированного дефицита природных киллеров, обусловленную мутацией в зоне центромеры 8-й хромосомы в локусе 8p11.23-q11.21. Один больной ребенок страдал тяжелым лимфопролиферативным синдромом, вызванным EBV, а двое других — рецидивирующими вирусными инфекциями дыхательной системы. Впоследствии в семье выявлено еще 4 случая такого иммунодефицита. Генеалогический анализ позволил установить аутосомно рецессивный тип наследования болезни.
В тот же год вышла еще одна статья C. Eidenschenk и соавторов (2006b), в которой описана особая форма семейного дефицита природных киллеров, который ассоциировался с нарушенными механизмами ИЛ-2- и ИЛ-15-зависимой выносливости лимфоцитов, в связи с чем у больных на фоне персистирующего низкого количества NK-клеток периодически отмечались эпизоды лимфопении.
G. Kabalak и соавторы (2010) описали генетически детерминированный дефицит ILT6 природных киллеров, ген которого располагается в локусе 19q13, и продемонстрировали ассоциацию этого иммунодефицита с синдромом Шегрена.
C. Gattazzo и соавторы (2010) установили, что у пациентов с лимфопролиферативным синдромом из больших гранулярных лимфоцитов (n=15) киллингингибирующий рецептор KIR3DL1 экспрессируется только в 13% случаев по сравнению с 90% в контрольной группе. Такие отличия связаны с аномально высоким метилированием промотера гена KIR3DL1, в связи с чем природные киллеры были лишены супрессивных сигналов, обычно ограничивающих их активацию и пролиферацию.
M. Machaczka и соавторы (2012) описали развитие лимфомы Ходжкина EBV-этиологии у взрослого с биалельной мутацией в гене STXBP2, регулирующего экзоцитоз перфоринсодержащих гранул природных киллеров.
F. Bernard и соавторы (2004) открыли новую форму первоначального дефицита природных киллеров, ассоциированную с дисморфией лица, нейтропенией и задержкой внутриутробного развития. Один ребенок умер в возрасте 18 мес от тяжелой CMV-инфекции, а его младшая сестра оставалась здоровой в 5-летнем возрасте, несмотря на наличие иммунодефицита.
L. Gineau и соавторы (2012) описали 6 родственных пациентов с парциальным дефицитом MCM4, у которых отмечалось отсутствие субпопуляции природных киллеров с фенотипом CD56(dim) и нормальное количество NK CD56(bright) с пониженной пролиферативной активностью. Ген MCM4 кодирует геликазный комплекс МСМ2-7, необходимый для репликации дезоксирибонуклеиновой кислоты (ДНК). У этих больных была идентифицирована мутация в сайте сплайсинга, что приводило к смещению рамки считывания и возникновению двух новых кодонов инициации трансляции вместо одного первичного. Кроме изолированного дефицита природных киллеров, у больных имела место задержка роста и недостаточность надпочечников, а при исследовании фибробластов отмечались признаки геномной нестабильности. В дальнейшем J.P. Casey и соавторы (2012) описали еще 10 пациентов с таким иммунодефицитом. Обзор этой проблемы осуществили C.R. Hughes и соавторы (2012).
Вирусные инфекции
Рецидивирующие вирусные инфекции – своеобразная визитка изолированного дефицита природных киллеров.
MG. Brown и соавторы (2001) доказали приветственную роль киллингактивирующих рецепторов природных киллеров в защите от вирусов.
В клинической картине преобладают частые респираторные вирусные инфекции (Isaacs D. et al., 1981) и инфекции, вызванные альфа-герпесвирусами (Orange J.S., 2002), что связано с недавно открытой ролью NK-клеток в контроле вирусов. (Backstrom E. et al., 2003). Кроме того, S. Nandakumar и соавторы (2008) показали, что в случае HSV-1-инфекции природные киллеры выполняют своеобразную хелперную функцию в отношении цитотоксических Т-лимфоцитов, играющих важную роль в контроле над HSV-1.
Рецидивирующий орофациальный и генитальный герпес обнаруживают чаще всего (Sirianni M.C. et al., 1986; Cauda R. et al., 1989; Cheknev S.B. et al., 1994; Forbes L.R. et al., 2009; 2); Однако иногда герпесвирусные инфекции могут принимать тяжелое течение с поражением нервной системы и внутренних органов, в частности — в виде диссеминированной ветряной оспы, CMV-пневмонита и распространенной кожной HSV-1-инфекции. О таком случае сообщили C.A. Biron и соавторы (1989). Хотя пациент с вирусными инфекциями выжил благодаря интенсивной терапии, в дальнейшем он умер от тяжелой апластической анемии (Orange J.S., 2002).
F. Almerigogna и соавторы (2011) описали 5 детей с энцефалитом, вызванным HSV-1, у которых отмечался дефицит природных киллеров. У двух из них удалось идентифицировать мутацию в гене транскрипта CD16A.
R. Maia и соавторы (2011) показали, что ранний рецидив герпесвирусного энцефалита может быть обусловлен дефицитом NK-клеток.
A. Etzioni и соавторы (2005) сообщили о двухлетней девочке, у которой отмечалась роковая диссеминированная ветряная оспа и отсутствовали естественные киллеры в крови, хотя все остальные изученные показатели иммунитета находились в пределах диапазона нормы. Родители пациентки были родственниками, а дефицит NK-клеток отмечался с самого рождения, еще до момента первичного контакта с VZV.
L.D. Notarangelo, E. Mazzolari (2006) также сообщили о серии случаев тяжелых форм VZV-инфекции у детей с изолированным дефицитом NK-клеток.
M.T. Vossen и соавторы (2005) описали 5 детей с угрожающей жизни ветряной оспой, у которых отмечался глубокий дефицит природных киллеров. Кроме экзантемы, у больных имел место VZV-пневмонит, гепатит и энцефалит. Один ребенок умер от полиорганной недостаточности, в то время как 4 других выжили благодаря интенсивной терапии. В них закономерно отмечалась бактериальная суперинфекция — в крови обнаружили Haemophilus influenzae типа b, Streptococcus pneumoniae и бета-гемолитический стрептококк группы А. Бактериемию можно объяснить тяжелым общим состоянием детей, однако P. Garcia-Penarrubia и 9 .
В последнее время появились многие сообщения о развитии реактивированных инфекций, вызванных лимфотропными герпесвирусами, у пациентов с изолированным дефицитом природных киллеров. V. Bekiaris и соавторы (2009) продемонстрировали, что природные киллеры защищают вторичную лимфоидную ткань от CMV благодаря CD30-зависимому механизму, а T. Strowig и соавторы (2008) показали, что именно тонзиллярные NK-клеток ограничивают процесс трансформации В-лимфоцитов EBV путем продукции гамма-ИФН.
F. Bernard и соавторы (2004) описали двух сестер с семейным дефицитом природных киллеров, одна из которых умерла от тяжелой СМV-инфекции, а другая оставалась здоровой в возрасте 5 лет, несмотря на сохранение иммунодефицита.
N. Rai, N. Thakur (2013) описали врожденную CMV-инфекцию у новорожденного, вызванную дефицитом природных киллеров. Ранее подобную ассоциацию обнаружили R. Cauda и соавторы (1987) в небольшом исследовании (n=29).
EBV у пациентов с дефицитом природных киллеров может быть причиной лимфопролиферативных синдромов (Eidenschenk C. et al., 2006a), опухоли из гладких мышц в надпочечниках (Shaw R.K. et al., 2012), хронического моно. , 1989;Aoukaty A. et al., 2003), синдрома хронической усталости (Caligiuri M. et al., 1987), фатального инфекционного мононуклеоза (Fleisher G. et al., 1982), гемофагоцитарного лимфогиоC. , 1988; Qiang Q. et al., 2012). В частности, G. Fleisher и соавторы (1982) описали 3 пациентов с дефицитом природных киллеров, у которых отмечалась прогрессирующая EBV-инфекция, первый из которых умер от фатального инфекционного мононуклеоза в детстве, второй — от легочной недостаточности, обусловленной вирусным пневмо лет, а третий оставался живым с клиникой хронического мононуклеоза и частых острых респираторных вирусных инфекций.
Z.K. Ballas и соавторы (1990) описали пациентку с количественным дефицитом природных киллеров и экспансией клеток с фенотипом CD3+CD4–CD8–NKH1+. У нее отмечались рецидивирующие кондиломы, вульварная и цервикальная карцинома in situ, пульмональные инфильтраты и состояние гиперкоагуляции, обусловленные папилломавирусной инфекцией. Больная умерла в возрасте 30 лет, а ее сестра – в детстве от лейкоза. Кроме классических проявлений в виде кожных папиллом (Cac N.N., Ballas Z.K., 2006), кондилом, дисплазии и рака шейки матки, у таких больных папилломавирусы могут поражать дыхательную систему, в частности быть причиной гранулематоза легких и рецидивирующего папиломатоза2.
У пациентов с дефицитом природных киллеров может развиваться персистирующая лимфаденопатия, вызванная Т-лимфотропным вирусом человека III типа (human T-cell lymphotropic virus type III - HTLV-III) (Fontana L. et al., 1986).
G.E. Kaiko и соавторы (2010) отметили дефицит естественных киллеров у новорожденных с хронической инфекцией, вызванной респираторно-синцитиальным вирусом.
R. Gazit и соавторы (2006) на животной модели дефицита рецептора Ncr1 природных киллеров показали ассоциацию этого иммунодефицита с летальной инфекцией, вызванной вирусом гриппа. В дальнейшем A. Fox и соавторы (2012) выявили дефицит естественных киллеров у некоторых пациентов с тяжелыми формами гриппа, вызванного пандемическим штаммом H1N1. H. Nave и соавторы (2011) отметили, что уменьшение количества природных киллеров в крови, иногда встречающееся у пациентов с ожирением, оказывает неблагоприятное влияние на тяжесть гриппа, обусловленного штаммом H1N1.
Еще в 1983 г. J.F. Bukowski и соавторы продемонстрировали в эксперименте, что извлечение природных киллеров усиливает репродукцию вируса в гепатоцитах и усугубляет проявления вызванного этим агентом гепатита. N. Muhanna и соавторы (2011) на животной модели показали подавление развития фиброза печени, обусловленного вирусом гепатита С, при активации природных киллеров. Ранее J. Corado и соавторы (1997) продемонстрировали дисфункцию природных киллеров у многих пациентов с вирусным гепатитом С.B. Yoffe и соавторы (1986) показали сохранение репродуктивной активности вируса гепатита В у мужчин с постнекротическим циррозом при наличии дефицита природных киллеров и более благоприятное течение инфекции у иммунокомпетентных лиц.
Автоиммунные осложнения
У пациентов с дефицитом естественных киллеров часто возникают аутоиммунные осложнения, однако остается до конца не понятным механизм их развития. Считается, что именно хронические вирусные инфекции являются причиной срыва иммунной толерантности к антигенам собственного организма у таких больных. Кроме того, иммунорегуляторные нарушения, в частности цитокиновый дисбаланс и неспособность к адекватной реализации реакций спонтанной или антителозависимой клеточно-опосредованной цитотоксичности, также могут объяснить некоторые случаи автоиммунной патологии у пациентов с изолированным дефицитом природных киллеров (Wood S.M. et al.). О механизмах автоиммунитета при дефиците NK-клеток подробно говорится в обзоре, осуществленном N. Schleinitz и соавторами (2010).
Впервые развитие автоиммунного осложнения, а именно системной красной волчанки (СКВ), при дефиците природных киллеров показали P. Katz и соавторы (1982). В дальнейшем H. Matsuzaki и соавторы (1983) описали пациента с инсулинзависимым сахарным диабетом и болезнью Грейвса, у которого отсутствовали признаки антитело-зависимой клеточно-опосредованной цитотоксичности с участием природных киллеров, однако имели нормальное течение реакции спонтанной цитотоксичности, инфекций и видоизменяло клиническую картину болезни. Позже R.I. Fox и соавторы (1984) установили, что у всех 5 исследованных пациентов с ревматоидным артритом, которых лечили путем удаления лимфоцитов с помощью дренажа грудного лимфатического протока, резко уменьшено количество природных киллеров в лимфе, хотя содержание других субпопуляций лимфоцитов.
M.R. Green и соавторы (2005) продемонстрировали функциональный дефицит природных киллеров в семьях, в которых были случаи СКВ и ревматоидного артрита, заключавшегося в сниженной активности килинга мишеней и слабой продукции цитокинов. Авторы подчеркнули, что причиной аутоагрессии у таких пациентов может быть не только хроническая вирусная инфекция, но и иммунная дизрегуляция, обусловленная потерей регуляторных воздействий со стороны природных киллеров. Согласно этому, J. Villanueva и соавторы (2005) показали, что дисфункция природных киллеров является типичным признаком ювенильного ревматоидного артрита и синдрома гиперактивации макрофагов (n=20).
A. Abrahamov, M. Schlesinger (1990) описали количественный дефицит естественных киллеров у 16-летнего пациента, страдавшего апластической анемией с четырех лет. Ранее P. Gascon и соавторам (1986) удалось набрать целую группу пациентов с апластической анемией (n=43), у которых отмечался изолированный дефицит NK-клеток. Впервые об ассоциации дефицита природных киллеров и апластической анемии сообщили E.P. Pogorel'skaia и соавторы (1985). У больных с этим иммунодефицитом могут также случаться случаи alopecia areata (Lutz G. et al., 1981), первичного билиарного цирроза (James S.P., Jones E.A., 1985), автоиммунной нейтропении (Thyss A. et al., 1, t. et al., 1996), сахарного диабета 1-го типа (Akesson C. et al., 2010; Jobim M., et al., 2010), синдрома Шегрена (Kabalak G. et al., 2009), СКВ (Cederblad B. et al., 1998;Park Y.W. et al., 2009), автоиммунного лимфопролиферативного синдрома с развитием витилиго, псориатического артрита, тиреоидита, узловой эритемы, билатерального переднего склерита и синдрома Ш.2.
J. Hao и соавторы (2010) показали, что резидентные природные киллеры в центральной нервной системе угнетают Th17, предотвращая развитие автоиммунной патологии. Это может объяснить известную ассоциацию дефицита ILT6 и рассеянного склероза у людей, которую впервые продемонстрировали S. Koch и соавторы в 2005 г.
Синдром опухолевого роста
Онкологические осложнения у больных с дефицитом природных киллеров можно объяснить потерей непосредственного иммунного наблюдения за мутантными клетками, а также реактивацией некоторых лимфотропных вирусов с онкогенными свойствами, в частности EBV. Последний механизм определяет преобладание лимфопролиферативных опухолей в клинической картине заболевания.
H.W. Ziegler и соавторы (1981) впервые сообщили об ассоциации изолированного дефицита природных киллеров и злокачественных опухолей. Был изучен иммунный статус у 10 больных с хроническим лимфолейкозом по сравнению с таковым в контрольной группе. Установлено, что 9 из 10 пациентов не определяли активность природных киллеров, несмотря на повышенный уровень этих клеток в крови. При измерении этой активности в литических единицах в 1 мл крови оказалось, что цитотоксический потенциал NK-клеток был по крайней мере в 7 раз ниже, чем у контрольной группы. Лечение с помощью фибробластного ИФН и индуктора интерфероногенеза 1:С было эффективным у 2 больных с ранними проявлениями неоплазии, однако не привело к улучшению у других 3 пациентов с болезнью.
Впоследствии L.P. Ruco и соавторы (1983) описали глубокий дефицит естественных киллеров у всех 10 обследованных лиц с волосококлеточной лейкемией в небольшом контролируемом исследовании. Авторы также показали эффективность препаратов альфа-ИФН в таких случаях. Позже B.R. Smith и соавторы (1985) сообщили еще о 4 больных с волосатоклеточным лейкозом, у которых отмечался глубокий функциональный дефицит природных киллеров.
Y. Fujimiya и соавторы (1987) выявили количественный и функциональный дефицит природных киллеров у лиц с хроническим миелолейкозом и предложили новый метод лечения – клонирование и амплификацию NK-клеток. Ранее такую ассоциацию описали HW. Ziegler-Heitbrock и соавторы (1985).
C.M. Yang и соавторы (2002) сообщили о тайваньской девочке с дефицитом естественных киллеров, у которой с 3-летнего возраста отмечался атопический дерматит и рецидивирующий лабиальный герпес, а в 6 лет развилась лимфома Ходжкина.
Кроме того, о причинной роли дефицита природных киллеров сообщали при гемофагоцитарном лимфогистиоцитозе (Imashuku S. et al., 2002; Schneider E.M. et al., 2003; Horne A. et al., 2005), метастатической Hol. 2011), лимфоме Ходжкина (Machaczka M. et al., 2012), хроническом лимфолейкозе (Platsoucas C.D. et al., 1980), недифференцированном остром лейкозе (Fang X.C. et al., 2010). et al., 2010).
Аллергические осложнения
Аллергия не является характерным признаком дефицита природных киллеров, в отличие от многих других иммунодефицитов. Как уже отмечалось выше, C.M. Yang и соавторы (2002) описали рецидивирующий атопический дерматит у пациентки с верифицированным диагнозом дефицита природных киллеров. Иногда встречается рецидивирующая мультиформная эритема, ассоциированная с HSV-1 (Messina M.F., 2011). На примере животной модели G.E. Kaiko и соавторы (2010) продемонстрировали, как хронический бронхит, вызванный респираторно-синцитиальным вирусом, трансформируется в инфекционно-зависимую астму в условиях дефицита NK-клеток. В частности, происходит постепенное переключение на Th2-ответ под влиянием продуцируемого эпителиоцитами ИЛ-25, тогда как у здоровых мышей, зараженных респираторно-синцитиальным вирусом, иммунный ответ реализовывался преимущественно по Th1-типу без развития аллергических осложнений. У пациента с дефицитом молекулы CD122 природных киллеров M. Inaoka (2005) описал тяжелую реакцию гиперчувствительности к медикаменту по типу DIHS/DRESS (Drug-Induced Hypersensitivity Syndrome/Drug Reaction with Eosinophilia and Systemic Symptoms), (human herpes virus) 6-го типа (HHV-6).
Другие симптомы
О развитии дилатационной кардиомиопатии у пациентов с дефицитом природных киллеров впервые сообщили JL. Андерсон и соавторы (1982). T. Itagaki и соавторы (1988) подтвердили ассоциацию дефицита природных киллеров и дилатационной кардиомиопатии, в патогенезе которой рассматривается участие вирусных агентов. L.V. Doering и соавторы (1997) продемонстрировали, что показатели гемодинамики и психоэмоциональный стресс не влияют на цитотоксичность, опосредованную природными киллерами, у больных с ишемической и дилатационной кардиомиопатией.
D.D. Kahana и соавторы (2003) описали случай склерозирующего холангита, вызванного грибковой инфекцией (Trichosporon), у 8-летней девочки с изолированным дефицитом природных киллеров. H. Akiba и соавторы (2001) сообщили о развитии трихофитической гранулемы у 28-летнего пациента с дефицитом природных киллеров, у которого имел место СКВ. Грибковое осложнение возникло после назначения кортикостероидов с целью подавления активности аутоиммунной патологии. R. Krishnaraj, A. Svanborg (1993) описали ассоциацию дефицита природных киллеров с диссеминированным аспергиллезом, вызванным Aspergillus fumigatus.
Дефицит природных киллеров может быть причиной инфекционных осложнений ран после хирургических операций на щитовидной железе, как сообщили H. Cerwenka и соавторы (2001).
I.F. Kruglova (1995) продемонстрировала дефицит крупных гранулярных лимфоцитов у больных пульмональным туберкулезом. H. Schmitt и соавторы (1999) описали легочную инфекцию, вызванную Mycobacterium xenopi, у пациента с дефицитом природных киллеров. T. Wendland и соавторы (2000) сообщили о диссеминированной инфекции, вызванной Mycobacterium avium, при этом иммунодефиците, ранее проявлявшимся рецидивирующей VZV-инфекцией. M.A. Karalian (2009) сообщили о 117 больных с микст-инфекцией, вызванной вирусными (HSV-2, CMV, вирус гепатита С) и микобактериальными агентами, у которых отмечались признаки дефицита природных киллеров. N. Samileh и соавторы (2006) описали изолированный дефицит клеток NK у детей с лимфаденитом после введения БЦЖ.
T. Aoki и соавторы (1987) описали группу больных (n=23)с изолированным дефицитом природных киллеров, у которых отмечались случаи ремитирующей лихорадки в течение длительного срока (?6 мес) или признаки синдрома хронической усталости. Возможно развитие ювенильного рецидивирующего паротита (Lopez Pico E. et al., 2012).
Компенсация иммунодефицита
Дефицит естественных киллеров иногда бессимптомно, в связи с чем исследователей всегда интересовали механизмы компенсации иммунной недостаточности в таких случаях, поскольку полученные знания могли бы быть полезными при разработке иммунотерапевтических вмешательств пациентам с клинически манифестными формами болезни. T. Wendland и соавторы (2010) продемонстрировали, что в случае диссеминированной инфекции, вызванной Mycobacterium avium, дефицит природных киллеров частично компенсировался за счет усиления ответа со стороны альфа-/бета- и гамма-/дельта-Т-лимфоцитов. В то же время, сами природные киллеры могут компенсировать другие иммунодефициты. В частности, A. Ferrante и соавторы (1985) обнаружили повышенное количество NK-клеток у пациентов после спленэктомии, у которых не отмечалось инфекционных осложнений, и объяснили этим феноменом компенсированное состояние этих пациентов. Однако у некоторых пациентов после спленэктомии происходит срыв компенсаторных механизмов и истощение природных киллеров, что приводит к частым острым респираторным вирусным инфекциям, рецидивному генитальному герпесу и хроническому лимфолейкозу, как это продемонстрировали J. Demeter и соавторы (1990) в небольшом ретроспективе. ).
Трансплантология
O. Schofer и соавторы (1989) впервые убедительно продемонстрировали участие естественных киллеров в реакциях отторжения трансплантата. K. Hadaya и соавторы (2008) показали, что сниженная способность природных киллеров к продукции гамма-ИФН и повышение экспрессии рецепторов NKG2C и NKG2D были предикторами высокого риска развития реактивированной CMV-инфекции у реципиентов аллоговой почки. I. Nowak и соавторы (2012) продемонстрировали в контролируемом исследовании, что делеция 22-bp в гене KIR2DS4 естественных киллеров определяла неблагоприятный прогноз в случае пересадки аллогенной почки (n=283). R.E. Cirocco и соавторы (2007) установили, что дефицит килингингибирующего рецептора 2DL2 так же, как и недостаточность килингактивирующей молекулы 2DS2, ассоциированы с неудовлетворительной функцией трансплантата через 1 год после пересадки аллогенной почки (n=12).
Диагностика
Определение количества лимфоцитов с фенотипом CD3–CD16+CD56+ с помощью проточной цитофлуориметрии позволяет идентифицировать количественный дефицит природных киллеров во многих случаях (Whiteside T.L. et al., 1990; Gineau L. et al., 202. 202). В случае дефицита азурофильных гранул содержание CD3–CD16+CD56+ нормальное, однако резко уменьшено количество крупных гранулярных лимфоцитов, определяемых при цитологическом исследовании мазка крови (Kay N.E., Zarling J.M., 1984). Иногда отмечается дефицит природных киллеров исключительно в лимфе при нормальном содержании этих клеток в крови (Fox R.I. et al., 1984). Дефицит CD3–CD16+CD56+-клеток может быть обусловлен низкой экспрессией молекул CD16+ или CD56+. В таком случае содержание больших гранулярных лимфоцитов в крови остается нормальным. При изолированном дефиците CD16 параллельно отмечается фагоцитарная дисфункция, поскольку этот маркер экспрессируют нейтрофилы. Это может привести к изменению клинической картины за счет присоединения бактериальных или грибковых инфекций. B.N. Jaeger и соавторы (2012) продемонстрировали, что удаление нейтрофилов нарушает созревание, функционирование и гомеостаз природных киллеров. В то же время возможен дефицит молекулы СD16 только на фагоцитах при нормальной ее экспрессии на поверхности природных киллеров. В таком случае может развиваться изоиммунная нейтропения новорожденных (Huizinga T.W. et al., 1990).
В случае качественного дефицита природных киллеров имеет место нарушенная функциональная активность этих клеток при нормальном или даже повышенном их количестве в крови. Функцию природных киллеров оценивают по продукции цитокинов, адгезии к мишеням и активности килинга, измеряемой в литических единицах. Добавление ИЛ-2 (Fujimiya Y. et al., 1987) или альфа-ИФН (Komiyama A. et al., 1990) к культуре NK-клеток позволяет изучить секреторную функцию природных киллеров по интенсивности высвобождения цитокинов. Форбол міристат стимулирует преимущественно выработку ИЛ-1?, а фитогемагглютинин-М - гамма-ИФН и ИЛ-2 (Chang W.C. et al., 1989; 1991). Фитогемагглютинин применяют в дозе 10 мкг/мл; результат оценивают через 48 часов инкубации (Chang W.C. et al., 1989; 1991). Спонтанную цитотоксичность определяют по активности адгезии и лизиса клеток-мишеней К652 (Virelizier J.L., Griscelli C., 1981), а антителозависимую - Р815 (de Vries E. et al., 1996). Результат оценивают в 4-часовом тесте высвобождения 51Cr (Komiyama A. et al., 1990), по цитотоксичности одной клетки на покровном стекле, покрытом поли-L-лизином (Fontana L. et al., 1986), или процентом специфической цитотоксичности проточной цитометрии (Devlin L.A. et al., 2010) Экспрессию перфорина оценивают с помощью иммуноферментного анализа винтактных клетках, а также в природных киллерах, стимулированных альфа-ИФН, присоединенными к полиэтиленгликолю (Portales P. et al., 2003).
Классификация
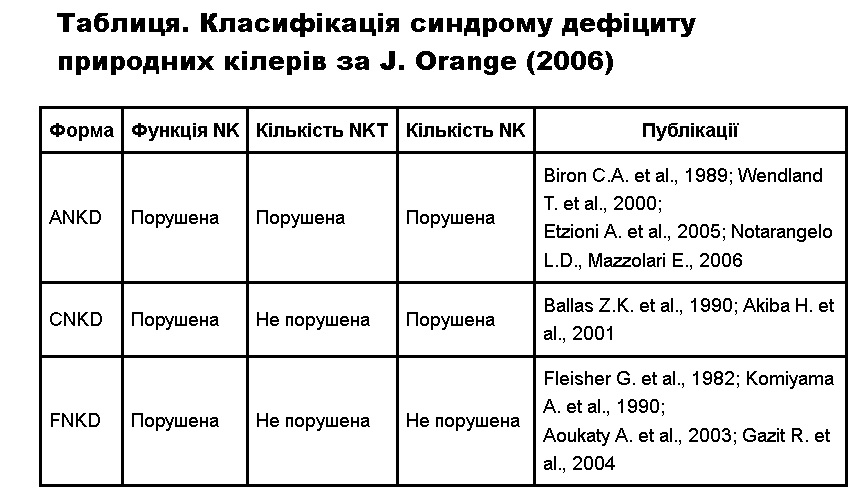
J. Orange (2006) предложил выделять три разновидности синдрома дефицита природных киллеров, в частности — абсолютный (Absolute natural killer cell deficiency — ANKD), классический (Classical natural killer cell deficiency — CNKD) и функциональный дефицит природных киллеров FNKD) (таблица).
Таблица. Классификация синдрома дефицита природных киллеров по J. Orange (2006)
Дефицит природных киллеров официально включен в авторитетную мировую базу данных On-line Mendelian Inheritance in Man (entry 609981). Международные эксперты предложили рабочий вариант критериев диагностики этого иммунодефицита (http://bioinf.uta.fi/xml/idr/ff/FF135.xml?style=pdf).
Дифференциальная диагностика
Необходимо проводить дифференциальный диагноз между первичным и вторичным дефицитом естественных киллеров.
Кроме того, существует несколько заболеваний иммунной системы с подобными клиническими проявлениями, которые следует иметь в виду при проведении дифференциальной диагностики. Речь идет о выборочных дефицитах альфа- и гамма-ИФН (Virelizier J.L., 1984), идиопатической Т-клеточной лимфопении, дефиците молекулы CD8 (Farhoudi A. et al., 2005), наследственной моноцитопении, дефицитах T. , 2012), ISG15 и NKT-клеток (Zeng W. et al., 2002).
Да, J.L. Virelizier, C. Griscelli (1981) описали мальчика в возрасте 4 года, страдавшего тяжелыми и частыми бактериальными (стафилококками, пневмококками, клебсиелами, моракселами) и вирусными (аденовирусами) инфекциями, у которого выявили выборочное нарушение продукции лейкоцитарных. Этот иммунодефицит приводил к вторичному снижению количества и функциональной активности природных киллеров. Назначение препарата альфа-ИФН с заместительной целью привело к восстановлению иммунного статуса.
C.A. Yang и соавторы (2012) сообщили о дисфункции Toll-like рецептора 3-го типа, обусловленной L412F-полиморфизмом гена ТLR-3, как причину рецидивирующего лабиального герпеса у людей. Ранее дефицит ТLR-3 был обнаружен у пациентов с височным HSV-1-энцефалитом, менингитом Молларе HSV-2-этиологии и рецидивирующим герпесвирусным стоматитом, которые могут встречаться у пациентов с изолированным дефицитом природных киллеров.
W. Zeng и соавторы (2002) сообщили о избирательной редукции субпопуляции NKT-клеток в костном мозге у пациентов с апластической анемией, часто развивающейся при дефиците NK-клеток. Кроме того, при недостаточности NKT-лимфоцитов отмечаются микобактериальные и вирусные инфекции, склонность к гиперергическим реакциям и целиакия.
J. Andersson и соавторы (1999) описали выборочный дефицит гамма-ИФН у 28-летней пациентки, у которой развилась тяжелая реактивированная EBV-инфекция с развитием интерстициального пневмонита, многоочагового лейкоэнцефалита и язв слизистой генитального тракта. Ремиссия болезни была достигнута за счет длительной заместительной терапии препаратом рекомбинантного гамма-ИФН. Лечение ацикловиром, преднизолоном и внутривенным (в/в) иммуноглобулином, проводившееся до этого момента, не приводило к кардинальному улучшению состояния пациентки.
D.C. Vinh и соавторы (2010) описали аутосомно-доминантную моноцитопению, приводившую к развитию рецидивирующих микобактериальных, вирусных, грибковых инфекций и миелодисплазии, что напоминало дефицит природных киллеров.
D. Bogunovic и соавторы (2012) идентифицировали наследственный дефицит молекулы ISG15 – ИФН-индуцибельного внутриклеточного протеина, задействованного в реакциях клеточного иммунитета. У этих пациентов отмечался вторичный дефицит природных киллеров, в том числе слабая способность к продукции гамма-ИФН. Однако в клинике развивались исключительно микобактериальные инфекции, а стойкость к вирусам была сохранена.
Дефицит молекулы CD8, экспрессируемый цитотоксическими Т-лимфоцитами, может быть причиной аналогичных рецидивных вирусных инфекций, отмечаемых при дефиците природных киллеров (Farhoudi A. et al., 2005).
Подобный клинический спектр также отмечается у пациентов с идиопатической Т-клеточной лимфопенией, диагноз которой устанавливают пациентам с устойчивым и глубоким уменьшением количества CD4+ Т-лимфоцитов в крови при отсутствии признаков инфекции, вызванной вирусом иммунодефицита человека (ВИЧ), и нормальными другими показателями (Sternfeld T. et al., 2005).
Дефицит природных киллеров и другие иммунодефициты
Если говорить о связи изолированного дефицита природных киллеров и других иммунодефицитных болезней, то можно выделить, по крайней мере, три разные клинические ситуации. Дефицит природных киллеров может быть составной частью других иммунодефицитов с более широким фенотипом, например некоторых недифференцированных клеточных или комбинированных иммунодефицитов. В частности, S. Pahwa и соавторы (1983) сообщили о тяжелой CMV-инфекции у 20-месячного пациента, проявлявшейся фебрильной температурой, рецидивирующей пневмонией, экзантемой, лимфаденопатией, язвами слизистой ротовой полости. В иммунном статусе отмечалась нейтропения, пониженные концентрации IgG и IgA в сыворотке крови, нарушена функциональная активность Т-, В-лимфоцитов и природных киллеров. На основании этих данных установлен диагноз недифференцированного комбинированного иммунодефицита. Пациент получил 4 курса лечения препаратом альфа-ИФН в дозе от 2 до 1 млн международных единиц (МЕ) в течение 10; 28; 80 и 67 суток соответственно. Гипериммунную плазму вводили каждые 3 нед. Весь курс терапии длился 9 мес. Удалось добиться компенсации иммунодефицита и устранения реактивированной CMV-инфекции. S. Ng и соавторы (2011) сообщили о дефиците природных киллеров у пациента с выраженной гипертрофией тимуса, у которого в дальнейшем S. Masuda (2011) установил диагноз синдрома Гуда.
В то же время, возможно сосуществование у одного пациента изолированного дефицита природных киллеров и другой иммунодефицитной болезни, в состав которой обычно не входит дефицит NK-клеток, что связано с высокой частотой этой малой иммунной дисфункции в человеческой популяции. Например, J. Toth и соавторы (1983) сообщили о сочетании дефицита природных киллеров и С1q-компонента комплемента в семье, в которой отмечали случаи СКВ-подобного синдрома. D.B. Bittleman и соавторы (1994) описали сосуществование тотального дефицита С2 и NK-клеток у пациента, страдавшего рецидивирующими инфекциями, перикардитом и гломерулонефритом, у которого впоследствии развились острая ревматическая лихорадка и СКВ. M.D. Witt и соавторы (2002) описали тяжелый некротический лимфаденит HSV-1 этиологии у пациента, у которого отмечалось сочетание общего вариабельного иммунодефицита и дефицита природных киллеров. V.R. Bonagura и соавторы (1989) сообщили о пациенте с общим вариабельным иммунодефицитом, у которого на фоне в/в иммуноглобулинотерапии развилась пневмония, вызванная Pneumocystis carinii. При дообследовании у этого больного обнаружили глубокий функциональный дефицит естественных киллеров, которым объяснили развитие осложнения, характерного для клеточных иммунодефицитов. M. Clerici и соавторы (1988) обнаружили дефицит естественных киллеров у 57-летнего пациента с общим вариабельным иммунодефицитом, у которого развилась тяжелая EBV-инфекция. B. Ostenstad и соавторы (1997) описали дефицит естественных киллеров у мальчика с гипер-IgM-синдромом, обусловленным мутацией в гене CD40L. Интересно, что количество NK-клеток повысилось после назначения препарата рекомбинантного гранулоцитарного колониестимулирующего фактора по поводу нейтропении. E.G. Davies и соавторы (1982) обнаружили дефицит естественных киллеров у двух младенцев с синдромом задержанного отпадения пуповины. Аскорбиновая кислота частично восстанавливала подвижность нейтрофилов, но не влияла на NK-клетки. T. Inaba и соавторы (1989) описали дефицит IgM и природных киллеров у пациента, у которого развилась стафилококковая пиодерма и фатальный инфекционный мононуклеоз.
И, наконец, возможное развитие вторичного дефицита природных киллеров у больного с первичным иммунодефицитом, в фенотип которого не входит это нарушение. N. Adachi и соавторы (1997) описали у пациента с болезнью Швахмана транзиторный количественный и качественный дефицит природных киллеров вследствие дефицита витамина Е. Гиповитаминоз развился в связи с длительным диспептическим синдромом, обусловленным недостаточностью поджелудочной железы. Количество и функция NK-клеток восстановились после назначения альфа-токоферола.
Вторичный дефицит природных киллеров
При диагностике вторичного дефицита природных киллеров обязательно выяснение причины иммуносупрессии, а устранение иммуносупрессивного фактора или спровоцированного им метаболического нарушения должно приводить к восстановлению количества или функции NK-клеток. Так T. Mori и соавторы (2000) описали дефицит естественных киллеров у пациента с нефротическим синдромом с минимальными изменениями во время ремиссии болезни Ходжкина. Лечение преднизолоном устранило дисфункцию почек и способствовало компенсации иммунодефицита. J.M. Quesada и соавторы (1995) выявили дефицит естественных киллеров у лиц, находящихся на гемодиализе. Непосредственной причиной вторичной иммуносупрессии была недостаточность кальцитриола, поскольку применение препарата обусловило нормализацию иммунного статуса. R.E. Pollock и соавторы (2000) продемонстрировали временное угнетение функции природных киллеров под влиянием хирургического стресса после операций по поводу сарком и других солидных опухолей. Иммуносупрессия длилась около 18 ч и самоустранялась, если не приводила к инфекционным осложнениям. S.J. Oosterling и соавторы (2006) показали эффективность периоперативной терапии альфа-ИФН у хирургических больных для компенсации иммунодефицита, связанного с оперативным вмешательством. К причинам дефицита природных киллеров можно отнести железодефицитную анемию (Santos P.C., Falcao R.P., 1990), обширную талассемию (Akbar A.N. et al., 1986), шистосомоз (Gastl G.A., et. al., 1985), почечную недостаточность (Chen X.F., Wang P.X., 1989), анемию Фанкони (Hersey P. et al., 1982), тяжелую депрессию (Zorrilla E.P. et al., 2001), алкоголизм (Nair. , 1990), прием глюкокортикоидов (Nair M.P., Schwartz S.A., 1984). Однако при вторичной иммуносупрессии обычно формируется иммунодефицит с более широким фенотипом, поскольку поражаются и другие субпопуляции лимфоцитов, что облегчает дифференциальную диагностику с первичными изолированными дефицитами природных киллеров.
Лечебные вмешательства
В лечении нуждаются лица с клинически манифестными формами болезни (Wood S.M. et al., 2011). Можно выделить по крайней мере два направления терапии – базисное лечение и терапия клинических проявлений (инфекционных, автоиммунных и онкологических) (Orange J.S., 2013).
Диета
J. Barone, J.R. Hebert (1988) показали, что полиненасыщенные жирные кислоты ингибируют активность природных киллеров путем индукции синтеза простагландинов с иммуносупрессивными свойствами. Эту особенность следует учитывать при выборе диеты пациентам с верифицированным диагнозом изолированного дефицита NK-клеток, особенно у больных с сосудистыми факторами риска. G. Ravaglia и соавторы (2000) изучили влияние микронутриентного статуса на функцию природных киллеров у лиц старше 90 лет. Количество NK-клеток положительно коррелировало с содержанием цинка (мужчины: r=0,573; p=0,007; женщины: r=0,373; p=0,031) и селена в сыворотке крови (женщины: r=0,409; p=0,018). В свою очередь, цитотоксическая активность природных киллеров у женщин положительно коррелировала с сывороточной концентрацией витамина Е и убихинона (р<0,05). Следовательно, коррекция нарушенного микронутритивного статуса может рассматриваться как рациональный компонент терапии у больных с дефицитом природных киллеров, особенно лиц старческого возраста. E. Tapazoglou и соавторы (1985) на примере двух добровольцев показали, как извлечение цинка из рациона приводило к развитию дефицита природных киллеров, устранявшегося после восстановления полноценного питания. J.I. Allen и соавторы (1983) впервые описали подавление функции естественных киллеров при дефиците цинка у человека.
Базисная терапия
Природные и рекомбинантные альфа-ИФН показали эффективность при дефиците природных киллеров в исследованиях in vitro и in vivo. В частности, V.R. Bonagura и соавторы (1989) показали, что добавление альфа-ИФН в дозе 800 МЕ/мл в культуру природных киллеров у пациента с качественным иммунодефицитом приводило к постепенному росту функциональной активности этих клеток с 9,2•7,7% до 17,2• 14,9%. A.P. Kelly и соавторы (1984) провели двойное слепое плацебо-контролируемое исследование, в котором показали восстановление нарушенной функциональной активности природных киллеров у реципиентов аллогенной почки под влиянием рекомбинантной терапии альфа-ИФН, что обеспечивало профилактический эффект в отношении CMV-инфекции. W.R. Frederick и соавторы (1985) устранили индуцированное ВИЧ угнетение функции природных киллеров с помощью альфа-ИФН, обеспечив профилактический эффект эпизодов реактивации CMV (n=19). D.M. See, JG. Tilles (1996) провели двойное слепое плацебо-контролируемое исследование эффективности альфа2а-ИФН у пациентов с синдромом хронической усталости, у которых отмечался изолированный дефицит природных киллеров или сочетанное иммунное нарушение, дополнительно включавшее дисфункцию Т-клеток (n=26). Терапия продолжалась 12 нед. Продемонстрировано выраженное повышение активности природных киллеров – с 35,1±11,7 до 91,5±22,7 литических единиц (р<0,01) и повышение качества жизни пациентов, которые оценивали по критериям QOL. P. Portales и соавторы (2003) продемонстрировали, что альфа-ИФН восстанавливает вызванное ВИЧ угнетение экспрессии перфорина в природных киллерах in vivo. K. Shiozawa, S. Shiozawa (2006) продемонстрировали эффективность альфа-ИФН в низкой (450 МЕ/сут) дозе путем перорального применения у пациентов с аномально малым количеством природных киллеров в крови, страдавших болезнью Шегрена. N.N. Cac, Z.K. Ballas (2006) успешно применили препарат альфа-ИФН у пациента с дефицитом природных киллеров, проявлявшийся рецидивирующими папилломами.
Если говорить о механизме действия препаратов альфа-ИФН в случае этого иммунодефицита, то, кроме прямого противовирусного эффекта и непосредственной заместительной функции, это лекарство усиливает секреторную активность природных киллеров, увеличивает количество цитоплазматических гранул, потенцирует адгезию и киллинг мишеней, однако на проли степени.
Иногда происходит резистентность к альфа-ИФН, впервые описанная G. Fleisher и соавторами в 1982 г., а в дальнейшем основательно охарактеризована Y. Fujimiya и соавторами (1986), что требует проведения альтернативных иммунотерапевтических вмешательств.
Препараты бета- и гамма-ИФН также эффективны в лечении пациентов с дефицитом природных киллеров (Ziegler H.W. et al., 1981; Hersey P. et al., 1982; Bandyopadhyay S. et al., 1987), однако имеют меньшую доказательство. чем альфа-ИФН. Эти лекарственные средства, особенно препараты гамма-ИФН, могут быть испытаны у пациентов с резистентностью к терапии альфа-ИФН. Y. Shi, C.T. Lutz (2002) показали, что гамма-ИФН обеспечивает контроль над В-лимфоцитами, трансформированными под влиянием EBV, и этот эффект опосредован цитотоксическими Т-клетками, что может быть эффективным компенсаторным механизмом при дефиците природных киллеров. T. Fujisaki и соавторы (1993) продемонстрировали терапевтический эффект препарата рекомбинантного гамма-ИФН при тяжелой хронической активной EBV-инфекции в небольшом контролируемом исследовании.
Препараты рекомбинантного ИЛ-2 также показали эффективность лечения дефицита природных киллеров in vitro и in vivo. Кроме непосредственного заместительного действия и опосредованного усиления функции природных киллеров, ИЛ-2 обеспечивает выраженный пролиферативный эффект, который может быть полезным при количественных нарушениях. Однако высокая стоимость и частое развитие выраженного гриппоподобного синдрома ограничивают клиническое применение этого лекарства. A.H. Rook и соавторы (1983) продемонстрировали устранение депрессии природных киллеров и рост цитотоксического потенциала этих клеток в отношении инфицированных CMV лимфоцитов у пациентов с синдромом приобретенного иммунодефицита (n=6) под влиянием рекомбинантного ИЛ-2. M. Caligiuri и соавторы (1987) апробировали рекомбинантный ИЛ-2 у пациентов с дефицитом естественных киллеров, проявлявшихся синдромом хронической усталости EBV-этиологии (n=41). Продемонстрировано восстановление способности NK-клеток к килингу К562, хотя цитотоксический эффект в отношении EBV-инфицированных В-лимфоцитов оставался несколько ослабленным. A. Plebani и соавторы (2000) сообщили о полном восстановлении функции природных киллеров у пациента с синдромом Грисцели под влиянием препарата ИЛ-2. Согласно этому, N.E. Kay, J. Zarling (1987) с помощью рекомбинантного ИЛ-2 достигли восстановления функциональной активности NK-клеток у больного с изолированным дефицитом естественных киллеров, у которого развилась В-клеточная лейкемия. S. Specter и соавторы (1989) показали, что с помощью ИЛ-2 можно как устранить, так и предотвратить супрессию природных киллеров при употреблении дельта-9-тетрагидроканабиола. E. Lotzova и соавторы (1991) достигли генерации цитотоксических природных киллеров в крови и костном мозге пациентов с острым миелолейкозом при длительной рекомбинантной инфузии ИЛ-2. H.N. Lode и соавторы (1998) продемонстрировали опосредованную природными киллерами эрадикацию метастазов нейробластомы в костном мозге при проведении терапии с использованием ИЛ-2, связанным с помощью методов генной инженерии с антисиалоганглиозидными антителами GD-2. N.J. Meropol и соавторы (1996) показали, что ежедневные подкожные инъекции рекомбинантного ИЛ-2 в низкой дозе стимулируют природные киллеры in vivo без существенной токсичности. A. Toren и соавторы (2000) осуществляли успешную терапию средними и высокими дозами рекомбинантного ИЛ-2 в виде подкожных инъекций в течение двух лет после аутологической пересадки костного мозга у 5 детей с IV стадией нейробластомы. Отмечалось восстановление ранее нарушенного количества и функциональной активности природных киллеров, в частности, их способности уничтожать K562 и клетки нейробластомы. У одного пациента, которому преждевременно отменили иммунотерапию в связи с развитием энцефалопатии, случился рецидив опухоли.
Существуют единичные сообщения об эффективности ридостина (Cheknev S.B. et al., 1994), лентинана (Aoki T. et al., 1987) и тимопоэтина (Fiorilli M. et al., 1981) при дефиците естественных киллеров, однако эти данные в дополнительных исследованиях, прежде чем они могут быть рекомендованы к широкому применению в клинической практике.
Y. Fujimiya и соавторы (1987) разработали специальный метод заместительной терапии – клонирование и амплификацию природных киллеров, полученных от самого больного, в искусственных условиях при наличии альфа-ИФН и ИЛ-2. Ввиду технической сложности и высокой стоимости метода последний может быть апробирован только у некоторых иммуноскомпрометированных пациентов с тяжелыми клиническими симптомами в случае резистентности к доступным фармакотерапевтическим вмешательствам.
Трансплантация костного мозга представляется самым радикальным методом лечения тяжелых форм дефицита природных киллеров, однако пока не хватает сообщений и клинических исследований по этой проблеме. L.D. Notarangelo, E. Mazzolari (2006) провели успешную пересадку алогенного костного мозга ребенку в возрасте 4,5 лет с дефицитом природных киллеров, проявлявшимся тяжелой рецидивирующей VZV-инфекцией. Вместо этого C.A. Biron и соавторы (1989) показали смерть пациента с дефицитом естественных киллеров от осложнений после трансплантации по поводу рефрактерной апластической анемии. Ранее иммунодефицит манифестировал герпесвирусными инфекциями (HSV-1, VZV, CMV).
Другие терапевтические вмешательства
Иммуноглобулин в дозе 500 мг/кг массы тела в/в ингибирует функциональную активность природных киллеров, как это продемонстрировали C.B. Coulam, C. Goodman (2000) в группе беременных с угрозой преждевременных родов, что следует учитывать при планировании рациональной иммунотерапии, особенно у пациентов с сочетанием дефицита природных киллеров и гипо- или дисзимоглобулинемии, требующих применения иммуноглобулина в/в для компенсации недостатка . В то же время применение иммуноглобулина в/в способствует уменьшению рецидивов генитального герпеса (Masci S. et al., 1995). Более того, R.W. Finberg и соавторы (1992) продемонстрировали, что высокодозовая терапия в/в иммуноглобулином (2 г/кг) повышает количество и усиливает активность естественных киллеров человека, поэтому вопрос о целесообразности иммуноглобулинотерапии должен рассматриваться отдельно в каждом конкретном случае.
Поскольку даже низкодозовая терапия пероральными контрацептивами ингибирует активность природных киллеров (Baker D.A. et al., 1989), женщинам с дефицитом NK-клеток следует рекомендовать другие способы контрацепции.
Гипертермия усиливает функциональную активность природных киллеров in vivo (Frey B. et al., 2012), поэтому необходим рациональный подход к антипиретической терапии у пациентов с дефицитом NK-клеток.
Профилактическая противовирусная терапия с помощью ациклических аналогов гуанозина и имиквимода показана пациентам, в клинической картине которых преобладают рецидивирующие вирусные инфекции. В частности, такой подход с успехом применяется в Baylor College of Medicine (Orange J.S., 2013). C.A. Biron и соавторы (1989) успешно применяли у пациента с дефицитом природных киллеров длительную профилактическую терапию ацикловиром, за счет которой удавалось сдерживать проявления HSV-1- и VZV-инфекций, однако впоследствии больной умер от апластической анемии аутоиммунного происхождения, базисной терапии иммунодефицита
Вакцинация против HSV-1, VZV и папилломавирусов за счет индукции специфических цитотоксических Т-лимфоцитов может способствовать компенсации пониженной резистентности к указанным вирусным агентам у некоторых пациентов с дефицитом природных киллеров (Orange J.S., 2013). Однако при подобном иммунодефиците описаны случаи диссеминированной вакцина-социированной ветряной оспы (Banovic T. et al., 2011), что ставит под сомнение безопасность применения живых аттенуированных противовирусных вакцин у пациентов с дефицитом NK-клеток. Вакцинация БЦЖ противопоказана (de Vries E. et al., 1996; Samileh N. et al., 2006).
При бактериальных и грибковых инфекциях у пациентов с дефицитом природных киллеров показано дополнительное применение антимикробных химиопрепаратов. Лечение должно быть агрессивным, как и при других иммунодефицитах (Orange J.S., 2013). M.T. Vossen и соавторы (2005) доложили об успешном лечении бактериальных суперинфекций как осложнений тяжелой ветряной оспы у детей с дефицитом природных киллеров путем инфузионной антибиотикотерапии. H. Schmitt и соавторы (1999) сообщили об эффективном лечении легочной инфекции, вызванной Mycobacterium xenopi, с помощью комбинированной терапии с применением кларитромицина, рифабутина и спарфлоксацина у 36-летнего пациента с изолированным дефицитом естественных киллеров. T. Wendland и соавторы (2000) показали неэффективность стандартной антимикробной терапии у пациента с дефицитом естественных киллеров, у которого развилась фатальная диссеминированная инфекция, вызванная Mycobacterium avium. H. Akiba и соавторы (2001) вылечили тяжелую грибковую инфекцию, обусловленную Trichophyton rubrum, у 28-летнего пациента с дефицитом природных киллеров с помощью итраконазола, причем терапевтический эффект был достигнут только при концентрации препарата в сыворотке крови на уровне 198 . в очагах поражения – 210 нг/мл.
A. Kreutzman и соавторы (2012) выяснили, что отсутствие таких генов килингингибирующих рецепторов природных киллеров как KIR2DL5A (p=0,0489), 2DL5B (p=0,030) и 2DL5all (p=0,0272) было связано с лучшей реакцией на дазатинин, противоопухолевое средство группы ингибиторов тирозинкиназы, у больных с хроническим миелолейкозом (n=191). Подобная связь отмечена также в отношении двух килингактивирующих молекул — 2DS1 (p=0,061) и 2DS2 (p=0,071).
Прогноз
Течение дефицита природных киллеров непредсказуемо (Wood S.M. et al., 2011). Некоторые ранее здоровые пациенты умирали только от одного эпизода инфекции, другие болели длительное время без развития летальных осложнений (Vossen M.T. et al., 2005). Даже ближайшие родственники с одинаковым иммунодефицитом могут иметь принципиально разное течение болезни – от асимптомного до фатального (Bernard F. et al., 2004), что свидетельствует о роли индивидуального опыта контактов с инфекционными агентами и влиянии дополнительных факторов среды (Eidenschenk C. et al.). , 2006a). Абсолютный дефицит природных киллеров имеет более худший прогноз, чем классический и функциональный. В первом случае в детстве или молодом возрасте умерли 5 из 10 пациентов, о которых сообщили недавно, причем один из них выжил только благодаря трансплантации костного мозга. Во втором случае погибли только 3 из 10 больных (Orange J.S., 2013). Назначение иммуносупрессантов резко ухудшает прогноз, как это продемонстрировали H. Akiba и соавторы (2001), однако оказание медицинской помощи (и особенно адекватных иммунотерапевтических вмешательств) улучшает последствия болезни в клинически манифестных случаях (Notarangelo L.D., Mazzolari E., 2006).
Клинический случай 1
Больная Ш., 1965 года. Поступила в нейрохирургическое отделение с жалобами на приступы потери сознания, слабость в мышцах, головокружение, нарушение походки. Считает себя больной с 1992 г., когда неожиданно развился гемипарез справа. В 1994 г. произошло новое нападение, сопровождавшееся углублением двигательных расстройств в правых конечностях, моторной афазией, атаксией. В дальнейшем подобные приступы регулярно повторялись с нарастанием неврологического дефицита. Около 5 лет назад присоединились пароксизмы потери сознания с судорогами в мышцах конечностей, которые впоследствии приобрели полиморфный характер. В молодости перенесла острую ревматическую лихорадку.
При объективном осмотре обнаружено ретикулярное ливедо на коже (рис. 1). Сердце, легкие, органы брюшной полости, почки без особенностей. В неврологическом статусе отмечается понижение интеллекта до уровня деменции, нарушена критика к собственному состоянию, эйфоризация. Насильственный смех и плач. Мелкоразмашистый горизонтальный нистагм в обе стороны. Дизартрия, дисфагия, дисфония. Девиация кончика языка вправо при выдвижении. Тетрапарез с преимущественным поражением левых конечностей. Сухожильные и периостальные рефлексы повышены, вызываются из расширенных рефлексогенных зон. Положительные симптомы Бабинского, Штрюмпеля, Россолимо. Отмечаются субкортикальные рефлексы и симптом Маринеску – Радовичи. Выполняет координаторные пробы с промахом и интенционным тремором. Нарушение функции стояния и походки.
По поводу пароксизмов утраты сознания выполнена электроэнцефалография, с помощью которой удалось идентифицировать разряды обостренных медленных и острых потенциалов в левой височной доле с акцентом вперед (рис. 2).
В связи с ретикулярным ливедом проведено допплерографическое исследование сосудов — продемонстрировано диффузное утолщение комплекса интима-медиа с концентрическим сужением просвета сосудов (рис. 3).
Эхоэнцефалографическое исследование показало смещение расширенного М-эхо на 1,5-2 см, а также выраженность латеральных сигналов; пульсация достигала 40-50%.
На магнитно-резонансной томограмме (МРТ) визуализирована выраженная атрофия головного мозга с множественными очагами демиелинизации и энцефаломаляции в белом веществе больших полушарий (рис. 4).
Выполнена люмбальная пункция: ликвор прозрачный, белок – 0,099 г/л, реакция Нонне – Апельта – отрицательная, реакция Панди – положительная, глюкоза 3,9 ммоль/л, цитоз – 1 лимфоцит/мл.
Полимеразная цепная реакция (ПЦР) сыворотки крови и ликвора с видоспецифическими праймерами герпесвирусов оказалась отрицательными, однако отмечался аномальный индекс специфических IgG к VZV (1,301) и НHV-6 (0,159), тогда как индексы по HSV-1, CMV и иммуноглобулинов.
По данным общего анализа крови уровень лейкоцитов колебался в пределах 11,8-9,7-109/л.
Развернутые иммунологические исследования позволили выявить только одно нарушение – глубокий дефицит природных киллеров.
Иммунограммы неоднократно выполняли в двух независимых лабораториях (лаборатории нейроиммунологии Института нейрохирургии Национальной академии медицинских наук (НАМН) Украины и лаборатории иммунодиагностики Института иммунологии и аллергологии Национального медицинского университета имени А.А. Богомольца). Использовался метод проточной цитометрии с определением лимфоцитов с фенотипом CD16+ и CD3–CD16+CD56+. Количество клеток колебалось от 0,051 до 0,125-109/л при норме - 0,20-0,45-109/л. Большие гранулярные лимфоциты отмечались в мазках крови в следовых количествах. Тесты на антитела к двуспиральной ДНК, фосфолипидам, ревматоидный фактор, антинейтрофильные цитоплазматические антитела были отрицательными. Уровень антистрептолизина О (АСЛО) не превышал 200 МЕ/мл. С-реактивный белок – отрицательный.
После курса противовирусного лечения больной назначена длительная непрерывная иммунотерапия препаратом рекомбинантного альфа-ИФН,за счет чего удалось достичь сначала стабилизации, а впоследствии частичного регресса симптомов, однако с сохранением признаков инвалидности. Своевременная иммунодиагностика и иммунотерапия позволили избежать инвалидности в этом случае.
Это первое в мире сообщение об ассоциации дефицита природных киллеров с VZV-васкулитом и НHV-6-лейкоэнцефалитом, хотя о других тяжелых формах VZV-инфекции у таких больных сообщалось ранее (Etzioni A. et al., 2005; Vossen M.T. et al., 2005), а НHV-6 рассматривался в контексте реакций гиперчувствительности к лекарственным средствам (Inaoka M., 2005). Случаи VZV-васкулопатии (Мальцев Д.В., Климчук В.В., 2011) и HHV-6-лейкоэнцефалита (Мальцев Д.В., 2012) у пациентов с другими нарушениями в иммунитете описаны в предыдущих публикациях.
Клинический случай 2
Больная Л., 17 лет. Обратилась в Институт иммунологии и аллергологии с жалобами на кровоизлияния на коже, насильственные сокращения мышц лица и шеи, периодические отеки и боли в крупных суставах.
Из анамнеза выяснено, что в детстве часто болела ангинами. Неоднократно в мазках из задней стенки глотки бактериологически определялся бета-гемолитический стрептококк группы А. В возрасте 5 лет перенесла острую ревматическую лихорадку. После этого начал развиваться ревмокардит. На момент обследования имеет место недостаточность митрального клапана II степени с фиброзом створок по данным эхокадиографии. В возрасте 7 лет дебютировала малая хорея с типичным хореоатетозом, проявления которой купировались к 12-летнему возрасту. Однако впоследствии развился другой гиперкинез в виде тиков и вокализаций, отмечавшийся на момент обследования.
В 5 лет перенесла тяжелый инфекционный мононуклеоз, после которого, как стало известно сейчас, развилась хроническая активная EBV-инфекция, приведшая к новым осложнениям — сначала автоиммунной тромбоцитопенической пурпуре, затем — рецидивирующему артриту, а ныне — энцефаломиелиту.
Несколько раз перенесла эпизоды внебольничной бронхопневмонии, в том числе последний раз в январе 2013 г. Получала интенсивную терапию в отделении реанимации по месту жительства в связи с дыхательной недостаточностью ІІ степени. Там же выполнена пункция костного мозга, патология не обнаружена.
На МРТ отмечаются множественные очаги некроза и демиелинизации в белом веществе полушарий головного мозга и грудном отделе спинного мозга.
Путем ПЦР сыворотки крови в двух лабораториях (лаборатории нейроиммунологии и лаборатории нейробиохимии Института нейрохирургии НАМН Украины) обнаружена ДНК EBV. Резко повышена концентрация в сыворотке крови IgG в VCA EBV.
При иммунологическом обследовании обнаружено понижение концентрации С3-компонента комплемента и малое количество природных киллеров. Тесты повторялись трижды в интервале с июля 2012 по февраль 2013 г. Удельный вес природных киллеров, определяемый в различных лабораториях методом проточной цитометрии по содержанию лимфоцитов с фенотипом CD3–CD56+ и CD3–CD16+CD56+, когда % (при норме – 8–17%). Хотя природные киллеры обладают противобактериальными эффектами, коковая инфекция не слишком характерна для этого иммунодефицита, однако типична для дефицита компонентов комплемента, особенно учитывая развитие автоиммунных осложнений (хореи, ревмокардита и Pediatric Autoimmune Neuropsychiatric Disorders Associated with Strepto). Хроническая активная EBV-инфекция с соответствующими автоиммунными осложнениями, а также с поражением мозга, типична именно для дефицита NK-клеток.
Это первое в мире сообщение о сочетании дефицита С3 и природных киллеров. Ранее были опубликованы данные по сосуществованию дефицита NK-клеток и компонентов С1 (Toth J. et al., 1983) и С2 (Bittleman D.B. et al., 1994).
Выводы
Изолированный дефицит природных киллеров – распространенная и потенциально курабельная иммунная дисфункция, которая является причиной рецидивирующей вирусной инфекции, автоиммунных и онкологических осложнений и некоторых дополнительных симптомов у людей. Это фенотип ряда генетически детерминированных заболеваний иммунной системы и реже — вторичной иммуносупрессии.
Течение дефицита NK-клеток варьирует от бессимптомного до тяжелой, угрожающей жизни. Отмечается гетерогенность проявлений даже в пределах одной семьи. Опасность связана с атипично тяжелым течением вирусных инфекций, особенно вызванных альфа-герпесвирусами, а также EBV, онкологическими осложнениями, которые являются основной причиной преждевременной смерти таких пациентов, а также аутоиммунных процессов, оказывающих выраженный инвалидизирующий эффект.
Рациональная иммунодиагностика и иммунотерапия, а при необходимости и профилактическое противовирусное лечение, помогают компенсировать проявления иммунодефицита и должны быть как можно шире внедрены в клиническую практику. В то же время, существует потребность в дополнительных исследованиях по иммунотерапии изолированного дефицита природных киллеров.
Список использованной литературы:
Мальцев Д.В. (2009) Клинический полиморфизм семейного дефицита больших гранулярных лимфоцитов. Клиническая иммунология, аллергология, инфектология, 3(22): 36–40.
Мальцев Д.В. (2012) Прогресуюча мультифокальна лейкоенцефалопатія, асоційована з вірусом герпесу людини 6-го типу. Укр. мед. часопис, 1(87): 136–142 (http://www.umj.com.ua/article/20513).
Мальцев Д.В., Климчук В.В. (2011) Труднощі у діагностиці неврологічних ускладнень VZV-інфекції: випадки з клінічної практики. Укр. мед. часопис, 5(85): 122–127 (http://www.umj.com.ua/article/12523).
Abrahamov A., Schlesinger M. (1990) Absence of natural killer cells in a child with pure red blood cell aplasia. Am. J. Hematol., 35(4): 258–262.
Adachi N., Migita M., Ohta T. et al. (1997) Depressed natural killer cell activity due to decreased natural killer cell population in a vitamin E-deficient patient with Shwachman syndrome: reversible natural killer cell abnormality by alpha-tocopherol supplementation. Eur. J. Pediatr., 156(6): 444–448.
Akbar A.N., Fitzgerald-Bocarsly P.A., de Sousa M. et al. (1986) Decreased natural killer activity in thalassemia major: a possible consequence of iron overload. J. Immunol., 136(5): 1635–1640.
Akesson C., Uvebrant K., Oderup C. et al. (2010) Altered natural killer (NK) cell frequency and phenotype in latent autoimmune diabetes in adults (LADA) prior to insulin deficiency. Clin. Exp. Immunol., 161(1): 48–56.
Akiba H., Motoki Y., Satoh M. et al. (2001) Recalcitrant trichophytic granuloma associated with NK-cell deficiency in a SLE patient treated with corticosteroid. Eur. J. Dermatol., 11(1): 58–62.
Allen J.I., Perri R.T., McClain C.J. et al. (1983) Alterations in human natural killer cell activity and monocyte cytotoxicity induced by zinc deficiency. J. Lab. Clin. Med., 102(4): 577–589.
Almerigogna F., Fassio F., Giudizi M.G. et al. (2011) Natural killer cell deficiencies in a consecutive series of children with herpetic encephalitis. Int. J. Immunopathol. Pharmacol., 24(1): 231–238.
Anderson J.L., Carlquist J.F., Hammond E.H. (1982) Deficient natural killer cell activity in patients with idiopathic dilated cardiomyopathy. Lancet, 2(8308): 1124–1127.
Andersson J., Isberg B., Christensson B. et al. (1999) Interferon gamma (IFN-gamma) deficiency in generalized Epstein-Barr virus infection with interstitial lymphoid and granulomatous pneumonia, focal cerebral lesions, and genital ulcers: remission following IFN-gamma substitution therapy. Clin. Infect. Dis., 28(5): 1036–1042.
Aoki T., Usuda Y., Miyakoshi H. et al. (1987) Low natural killer syndrome: clinical and immunologic features. Nat. Immun. Cell. Growth. Regul., 6(3): 116–128.
Aoukaty A., Lee I.F., Wu J., Tan R. (2003) Chronic active Epstein-Barr virus infection associated with low expression of leukocyte-associated immunoglobulin-like receptor-1 (LAIR-1) on natural killer cells. J. Clin. Immunol., 23(2): 141–145.
Backstrom E., Chambers B.J., Ho E.L. et al. (2003) Natural killer cell-mediated lysis of dorsal root ganglia neurons via RAE1/NKG2D interactions. Eur. J. Immunol., 33(1): 92-100.
Baker D.A., Salvatore W., Milch P.O. (1989) Effect of low-dose oral contraceptives on natural killer cell activity. Contraception, 39(1): 119–124.
Ballas Z.K., Turner J.M., Turner D.A. et al. (1990) A patient with simultaneous absence of «classical» natural killer cells (CD3—, CD16+, and NKH1+) and expansion of CD3+, CD4–, CD8–, NKH1+ subset. J. Allergy Clin. Immunol., 85(2): 453–459.
Bandyopadhyay S., Miller D.S., Matsumoto-Kobayashi M. et al. (1987) Effects of interferons and interleukin 2 on natural killing of cytomegalovirus-infected fibroblasts. Clin. Exp. Immunol., 67(2): 372–382.
Banovic T., Yanilla M., Simmons R. et al. (2011) Disseminated varicella infection caused by varicella vaccine strain in a child with low invariant natural killer T cells and diminished CD1d expression. J. Infect. Dis., 204(12): 1893–1901.
Barone J., Hebert J.R. (1988) Dietary fat and natural killer cell activity. Med. Hypotheses, 25(4): 223–226.
Bekiaris V., Gaspal F., McConnell F.M. et al. (2009) NK cells protect secondary lymphoid tissue from cytomegalovirus via a CD30-dependent mechanism. Eur. J. Immunol., 39(10): 2800–2808.
Bernard F., Picard C., Cormier-Daire V. et al. (2004) A novel developmental and immunodeficiency syndrome associated with intrauterine growth retardation and a lack of natural killer cells. Pediatrics, 113(1 Pt 1): 136–141.
Biron C.A., Byron K.S., Sullivan J.L. (1989) Severe herpesvirus infections in an adolescent without natural killer cells. N. Engl. J. Med., 320(26): 1731–1735.
Bittleman D.B., Maves K.K., Bertolatus J.A. et al. (1994) Recurrent infections, pericarditis and renal disease in a patient with total C2 deficiency and decreased NK cell function consistent with acute rheumatic fever and systemic lupus erythematosus. Ann. Rheum. Dis., 53(4): 280–281.
Bogunovic D., Byun M., Durfee L.A. et al. (2012) Mycobacterial disease and impaired IFN-? immunity in humans with inherited ISG15 deficiency. Science, 337(6102): 1684–1688.
Bonagura V.R., Cunningham-Rundles S., Edwards B.L. et al. (1989) Common variable hypogammaglobulinemia, recurrent Pneumocystis carinii pneumonia on intravenous gamma-globulin therapy, and natural killer deficiency. Clin. Immunol. Immunopathol., 51(2): 216–231.
Bonagura V.R., Hatam L.J., Rosenthal D.W. et al. (2010) Recurrent respiratory papillomatosis: a complex defect in immune responsiveness to human papillomavirus-6 and -11. APMIS, 118(6–7): 455–470.
Bourin P., Mansour I., Doinel C. et al. (1993) Circadian rhythms of circulating NK cells in healthy and human immunodeficiency virus-infected men. Chronobiol. Int., 10(4): 298–305.
Brown M.G., Dokun A.O., Heusel J.W. et al. (2001) Vital involvement of a natural killer cell activation receptor in resistance to viral infection. Science, 292(5518): 934–937.
Bukowski J.F., Woda B.A., Habu S. et al. (1983) Natural killer cell depletion enhances virus synthesis and virus-induced hepatitis in vivo. J. Immunol., 131(3): 1531–1538.
Cac N.N., Ballas Z.K. (2006) Recalcitrant warts, associated with natural killer cell dysfunction, treated with systemic IFN-alpha. J. Allergy Clin. Immunol., 118(2): 526–528.
Caligiuri M., Murray C., Buchwald D. et al. (1987) Phenotypic and functional deficiency of natural killer cells in patients with chronic fatigue syndrome. J. Immunol., 139(10): 3306–3313.
Casey J.P., Nobbs M., McGettigan P. et al. (2012) Recessive mutations in MCM4/PRKDC cause a novel syndrome involving a primary immunodeficiency and a disorder of DNA repair. J. Med. Genet., 49(4): 242–245.
Cauda R., Laghi V., Tumbarello M. et al. (1989) Immunological alterations associated with recurrent herpes simplex genitalis. Clin. Immunol. Immunopathol., 51(2): 294–302.
Cauda R., Prasthofer E.F., Grossi C.E. et al. (1987) Congenital cytomegalovirus: immunological alterations. J. Med. Virol., 23(1): 41–49.
Cederblad B., Blomberg S., Vallin H. et al. (1998) Patients with systemic lupus erythematosus have reduced numbers of circulating natural interferon-alpha- producing cells. J. Autoimmun., 11(5): 465–470.
Cerwenka H., Wolf G., Mischinger H.J. et al. (2001) Natural killer cell deficiency and severe wound infection after thyroid surgery. Eur. J. Surg., 167(10): 792–794.
Chang W.C., Fujimiya Y., Casteel N., Pattengale P. (1989) Natural killer cell immunodeficiency in patients with chronic myelogenous leukemia. III. Defective interleukin-2 production by T-helper and natural killer cells. Int. J. Cancer, 43(4): 591–597.
Chang W.C., Hsiao M.H., Pattengale P.K. (1991) Natural killer cell immunodeficiency in patients with chronic myelogenous leukemia. IV. Interleukin-1 deficiency, gamma-interferon deficiency and the restorative effects of short-term culture in the presence of interleukin-2 on natural killer cytotoxicity, natural killer-target binding and production of natural killer cytotoxic factor. Nat. Immun. Cell Growth Regul., 10(2): 57–70.
Cheknev S.B., Mikovskaia O.I., Meshkova E.N. et al. (1994) The natural killer activity and indices of the interferon status of patients with recurrent genital herpes being treated with ridostin. Vopr. Virusol., 39(3): 125–128.
Chen X.F., Wang P.X. (1989) NK cell activity in patients with kidney insufficiency. Zhong Xi Yi Jie He Za Zhi, 9(7): 409–410.
Cirocco R.E., Mathew J.M., Burke G.W. 3rd et al. (2007) Killer cell immunoglobulin-like receptor polymorphisms in HLA-identical kidney transplant recipients: lack of 2DL2 and 2DS2 may be associated with poor graft function. Tissue Antigens, 69(1): 123–124.
Clerici M., Villa M.L., Mantovani M., Rugarli C. (1988) NK cell activity and monocyte dysfunctions in a patient with common variable hypogammaglobulinemia. J. Clin. Lab. Immunol., 27(3): 143–147.
Corado J., Toro F., Rivera H. et al. (1997) Impairment of natural killer (NK) cytotoxic activity in hepatitis C virus (HCV) infection. Clin. Exp. Immunol; 109: 451.
Coulam C.B., Goodman C. (2000) Increased pregnancy rates after IVF/ET with intravenous immunoglobulin treatment in women with elevated circulating C56+ cells. Early Pregnancy, 4(2): 90–98.
Davies E.G., Isaacs D., Levinsky R.J. (1982) Defective immune interferon production and natural killer activity associated with poor neutrophil mobility and delayed umbilical cord separation. Clin. Exp. Immunol., 50(2): 454–460.
de Vries E., Koene H.R., Vossen J.M. et al. (1996) Identification of an unusual Fc gamma receptor IIIa (CD16) on natural killer cells in a patient with recurrent infections. Blood, 88(8): 3022–3027.
Demeter J., Paloczi K., Lehoczky D., Benczur M. (1990) Observations on NK cells, K cells and on their function a long time after posttraumatic splenectomy. Int. Arch. Allergy Appl. Immunol., 92(3): 287–292.
Devlin L.A., Haughton D.J., Crockard A.D., Edgar J.D. (2010) Natural killer cell cytotoxicity in patients with recurrent herpes infections: diagnostic utility of a flow cytometric assay. J. Clin. Pathol., 63(3): 244–248.
Doering L.V., Vredevoe D.L., Woo M.A. et al. (1997) Predictors of natural killer cell-mediated cytotoxicity deficiency in advanced heart failure secondary to either ischemic or idiopathic dilated cardiomyopathy. Am. J. Cardiol., 80(2): 234–236.
Eidenschenk C., Dunne J., Jouanguy E. et al. (2006a) A novel primary immunodeficiency with specific natural-killer cell deficiency maps to the centromeric region of chromosome 8. Am. J. Hum. Genet., 78(4): 721–727.
Eidenschenk C., Jouanguy E., Alcais A. et al. (2006b) Familial NK cell deficiency associated with impaired IL-2- and IL-15-dependent survival of lymphocytes. J. Immunol., 177(12): 8835–8843.
Etzioni A., Eidenschenk C., Katz R. et al. (2005) Fatal varicella associated with selective natural killer cell deficiency. J. Pediatr., 146(3): 423–425.
Fang X.C., Liu H.L., Sun Z.M. et al. (2010) Expression and abscission of activated receptors and their ligands on/from NK cells in peripheral blood of patients with acute leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi, 18(2): 436–440.
Farhoudi A., Chavoshzadeh Z., Mir Saeid Ghazi B. et al. (2005) Recurrent infections and bilateral uveitis in a patient with CD8 deficiency. Iran. J. Allergy Asthma Immunol., 4(1): 43–45.
Ferrante A., Kiroff G.K., Goh D.H., Drew P.A. (1985) Elevated natural killer (NK) cell activity: a possible role in resistance to infection and malignancy in immunodeficient splenectomized patients. Med. Hypotheses, 16(2):133–146.
Finberg R.W., Newburger J.W., Mikati M.A. et al. (1992) Effect of high doses of intravenously administered immune globulin on natural killer cell activity in peripheral blood. J. Pediatr., 120(3): 376–380.
Fiorilli M., Sirianni M.C., Pandolfi F. et al. (1981) Improvement of natural killer activity and of T cells after thymopoietin pentapeptide therapy in a patient with severe combined immunodeficiency. Clin. Exp. Immunol., 45(2): 344-351.
Fleisher G., Starr S., Koven N. et al. (1982) A non-x-linked syndrome with susceptibility to severe Epstein-Barr virus infections. J. Pediatr., 100(5): 727–730.
Fontana L., Sirianni M.C., de Sanctis G. et al. (1986) Deficiency of natural killer activity, but not of natural killer binding, in patients with lymphoadenopathy syndrome positive for antibodies to HTLV-III. Immunobiology, 171(4–5): 425–435.
Forbes L.R., Paessler M., Orange J.S. (2009) CD16 deficiency discovered in a cohort of patients with severe herpesviral infections using a new diagnostic test. J. Allergy Clin. Immunol., 123(2 Suppl.): S67.
Fox A., Le N.M., Horby P. et al. (2012) Severe pandemic H1N1 2009 infection is associated with transient NK and T deficiency and aberrant CD8 responses. PLoS One, 7(2): e31535.
Fox R.I., Fong S., Tsoukas C., Vaughan J.H. (1984) Characterization of recirculating lymphocytes in rheumatoid arthritis patients: selective deficiency of natural killer cells in thoracic duct lymph. J. Immunol., 32(6): 2883–2887.
Frederick W.R., Epstein J.S., Gelmann E.P. et al. (1985) Viral infections and cell-mediated immunity in immunodeficient homosexual men with Kaposi’s sarcoma treated with human lymphoblastoid interferon. J. Infect. Dis., 152(1): 162–170.
Frey B., Weiss E.M., Rubner Y. et al. (2012) Old and new facts about hyperthermia-induced modulations of the immune system. Int. J. Hyperthermia, 28(6): 528–542.
Fujimiya Y., Bakke A., Chang W.C. et al. (1986) Natural killer-cell immunodeficiency in patients with chronic myelogenous leukemia. I. Analysis of the defect using the monoclonal antibodies HNK-1 (LEU-7) and B73.1. Int. J. Cancer, 37(5): 639–649.
Fujimiya Y., Chang W.C., Bakke A. et al. (1987) Natural killer (NK) cell immunodeficiency in patients with chronic myelogenous leukemia. II. Successful cloning and amplification of natural killer cells. Cancer Immunol. Immunother., 24(3): 213–220.
Fujisaki T., Nagafuchi S., Okamura T. (1993) Gamma-interferon for severe chronic active Epstein-Barr virus. Ann. Intern. Med., 118(6): 474–475.
Garcia-Penarrubia P., Koster F.T., Kelley R.O. et al. (1989) Antibacterial activity of human natural killer cells. J. Exp Med., 169(1): 99–113.
Gascon P., Zoumbos N., Young N. (1986) Analysis of natural killer cells in patients with aplastic anemia. Blood, 67(5): 1349–1355.
Gastl G.A., Feldmeier H., Kortmann C. et al. (1986) Human schistosomiasis: deficiency of large granular lymphocytes and indomethacin-sensitive suppression of natural killing. Scand. J. Immunol., 23(3): 319–325.
Gattazzo C., Teramo A., Miorin M. et al. (2010) Lack of expression of inhibitory KIR3DL1 receptor in patients with natural killer cell-type lymphoproliferative disease of granular lymphocytes. Haematologica, 95(10): 1722–1729.
Gazit R., Garty B.Z., Monselise Y. et al. (2004) Expression of KIR2DL1 on the entire NK cell population: a possible novel immunodeficiency syndrome. Blood., 103(5): 1965–1966.
Gazit R., Gruda R., Elboim M. et al. (2006) Lethal influenza infection in the absence of the natural killer cell receptor gene Ncr1. Nat. Immunol., 7(5): 517–523.
Gineau L., Cognet C., Kara N. et al. (2012) Partial MCM4 deficiency in patients with growth retardation, adrenal insufficiency, and natural killer cell deficiency. J. Clin. Invest., 122(3): 821–832.
Green M.R., Kennell A.S., Larche M.J. et al. (2005) Natural killer cell activity in families of patients with systemic lupus erythematosus: demonstration of a killing defect in patients. Clin. Exp. Immunol., 141(1): 165–173.
Hadaya K., de Rham C., Bandelier C. et al. (2008) Natural killer cell receptor repertoire and their ligands, and the risk of CMV infection after kidney transplantation. Am. J. Transplant., 8(12): 2674–2683.
Hao J., Liu R., Piao W. et al. (2010) Central nervous system (CNS)-resident natural killer cells suppress Th17 responses and CNS autoimmune pathology. J. Exp. Med., 207(9): 1907–1921.
Hersey P., Edwards A., Lewis R. et al. (1982) Deficient natural killer cell activity in a patient with Fanconi’s anaemia and squamous cell carcinoma. Association with defect in interferon release. Clin. Exp. Immunol., 48(1): 205–212.
Holtan S.G., Creedon D.J., Thompson M.A. et al. (2011) Expansion of CD16-negative natural killer cells in the peripheral blood of patients with metastatic melanoma. Clin. Dev. Immunol., 2011: 316314.
Horne A., Zheng C., Lorenz I. et al. (2005) Subtyping of natural killer cell cytotoxicity deficiencies in haemophagocytic lymphohistocytosis provides therapeutic guidance. Br. J. Haematol., 129(5): 658–666.
Hughes C.R., Guasti L., Meimaridou E. et al. (2012) MCM4 mutation causes adrenal failure, short stature, and natural killer cell deficiency in humans. J. Clin. Invest., 122(3): 814–820.
Hughes H.P., Kasper L.H., Little J., Dubey J.P. (1988) Absence of a role for natural killer cells in the control of acute infection by Toxoplasma gondii oocysts. Clin. Exp. Immunol., 72(3): 394–399.
Huizinga T.W., Kuijpers R.W., Kleijer M. et al. (1990) Maternal genomic neutrophil FcRIII deficiency leading to neonatal isoimmune neutropenia. Blood, 76(10): 1927–1932.
Hwang I., Zhang T., Scott J.M. et al. (2012) Identification of human NK cells that are deficient for signaling adaptor FcR? and specialized for antibody-dependent immune functions. Int. Immunol., 24(12): 793–802.
Imashuku S., Hyakuna N., Funabiki T. et al. (2002) Low natural killer activity and central nervous system disease as a high-risk prognostic indicator in young patients with hemophagocytic lymphohistiocytosis. Cancer, 94(11): 3023–3031.
Inaba T., Hanada R., Yaginuma A. et al. (1989) Fatal infectious mononucleosis with staphylococcal pyoderma in a girl with hereditary immunological dysregulations. Eur. J. Pediatr., 149(3): 177–178.
Inaoka M. (2005) Innate immunity and hypersensitivity syndrome. Toxicology, 209(2): 161–163.
Isaacs D., Clarke J.R., Tyrrell D.A. et al. (1981) Deficient production of leucocyte interferon (interferon-alpha) in vitro and in vivo in children with recurrent respiratory tract infections. Lancet, 2(8253): 950–952.
Itagaki T., Yamakawa K., Ono S. et al. (1988) Dilated cardiomyopathy associated with natural killer cell deficiency. Am. Heart. J., 115(6): 1326–1328.
Jaeger B.N., Donadieu J., Cognet C. et al. (2012) Neutrophil depletion impairs natural killer cell maturation, function, and homeostasis. J. Exp. Med., 209(3): 565–580.
James S.P., Jones E.A. (1985) Abnormal natural killer cytotoxicity in primary biliary cirrhosis: evidence for a functional deficiency of cytolytic effector cells. Gastroenterology, 89(1): 165–171.
Jawahar S., Moody C., Chan M. et al. (1996) Natural Killer (NK) cell deficiency associated with an epitope-deficient Fc receptor type IIIA (CD16-II). Clin. Exp. Immunol., 103(3): 408–413.
Jobim M., Chagastelles P., Salim P.H. et al. (2010) Association of killer cell immunoglobulin-like receptors and human leukocyte antigen-C genotypes in South Brazilian with type 1 diabetes. Hum. Immunol., 71(8): 799–803.
Joncas J.H., Ghibu C., Alfieri C. et al. (1989) Killer cell defect and persistent immunological abnormalities in two patients with chronic active Epstein-Barr virus infection. J. Med. Virol., 28(2): 110–117.
Kabalak G., Dobberstein S.B., Matthias T. et al. (2009) Association of immunoglobulin-like transcript 6 deficiency with Sjogren’s syndrome. Arthritis Rheum., 60(10): 2923–2925.
Kahana D.D., Cass O., Jessurun J. et al. (2003) Sclerosing cholangitis associated with trichosporon infection and natural killer cell deficiency in an 8-year-old girl. J. Pediatr. Gastroenterol. Nutr., 37(5): 620–623.
Kaiko G.E., Phipps S., Angkasekwinai P. et al. (2010) NK cell deficiency predisposes to viral-induced Th2-type allergic inflammation via epithelial-derived IL-25. J. Immunol., 185(8): 4681–4690.
Kaplan J., Shope T.C., Bollinger R.O., Smith J. (1982) Human newborns are deficient in natural killer activity. J. Clin. Immunol., 2(4): 350–355.
Karalian M.A. (2009) The influence of chronic viral infections on tuberculosis. Georgian Med. News, 168: 63–67.
Katz P., Zaytoun A.M., Lee J.H. Jr. et al. (1982) Abnormal natural killer cell activity in systemic lupus erythematosus: an intrinsic defect in the lytic event. J. Immunol., 129(5): 1966–1971.
Kay N.E., Zarling J. (1987) Restoration of impaired natural killer cell activity of B-chronic lymphocytic leukemia patients by recombinant interleukin-2. Am J. Hematol., 24(2): 161–167.
Kay N.E., Zarling J.M. (1984) Impaired natural killer activity in patients with chronic lymphocytic leukemia is associated with a deficiency of azurophilic cytoplasmic granules in putative NK cells. Blood, 63(2): 305–309.
Kelly A.P., Schooley R.T., Rubin R.H., Hirsch M.S. (1984) Effect of interferon alpha on natural killer cell cytotoxicity in kidney transplant recipients. Clin. Immunol. Immunopathol., 32(1): 20–28.
Koch S., Goedde R., Nigmatova V. et al. (2005) Association of multiple sclerosis with ILT6 deficiency. Genes Immun., 6(5): 445–447.
Komiyama A., Kawai H., Yabuhara A. et al. (1990) Natural killer cell immunodeficiency in siblings: defective killing in the absence of natural killercytotoxic factor activity in natural killer and lymphokine-activated killer cytotoxicities. Pediatrics, 85(3): 323–330.
Kreutzman A., Jaatinen T., Greco D. et al. (2012) Killer-cell immunoglobulin-like receptor gene profile predicts good molecular response to dasatinib therapy in chronic myeloid leukemia. Exp. Hematol., 40(11): 906–913.e1.
Krishnaraj R., Svanborg A. (1993) Low natural killer cell function in disseminated aspergillosis. Scand. J. Infect. Dis., 25(4): 537–541.
Kruglova I.F. (1995) The count of large granular lymphocytes in the blood and natural killing in tuberculosis. Lik. Sprava, (1–2): 81-84.
Lode H.N., Xiang R., Dreier T. et al. (1998) Natural killer cell-mediated eradication of neuroblastoma metastases to bone marrow by targeted interleukin-2 therapy. Blood, 91(5): 1706–1715.
Lopez Pico E., Vila Sexto L., Cacharron Carames T., Moreno Alvarez A. (2012) Juvenile recurrent parotitis and natural killer cells deficiency. An. Pediatr. (Barc.), 77(3): 216–217.
Lotzova E., Savary C.A., Schachner J.R. et al. (1991) Generation of cytotoxic NK cells in peripheral blood and bone marrow of patients with acute myelogenous leukemia after continuous infusion with recombinant interleukin-2. Am. J. Hematol., 37(2): 88–99.
Lutz G., Niedecken H., Bauer R., Kreysel H.W. (1988) Natural killer cell and cytotoxic/suppressor T cell deficiency in peripheral blood in subjects with alopecia areata. Australas. J. Dermatol., 29(1): 29–32.
Machaczka M., Klimkowska M., Chiang S. et al. (2012) Development of classical Hodgkin lymphoma in an adult with biallelic STXBP2 mutations. Haematologica, Oct. 25 [Epub ahead of print].
Maher K.J., Klimas N.G., Fletcher M.A. (2005) Chronic fatigue syndrome is associated with diminished intracellular perforin. Clin. Exp. Immunol., 142(3): 505–511.
Maia R., Gouveia C., Moreira A. et al. (2011) Early ‘’relapse’’ after herpetic encephalitis: extensive white matter lesions in an infant with interferon production deficit. J. Child Neurol., 26(3): 369–372.
Masci S., De Simone C., Famularo G. et al. (1995) Intravenous immunoglobulins suppress the recurrences of genital herpes simplex virus: a clinical and immunological study. Immunopharmacol. Immunotoxicol., 17(1): 33–47.
Masuda S. (2011) More on NK-cell and B-cell deficiency with a thymic mass. N. Engl. J. Med., 364(20): 1979–1980.
Matsuzaki H., Kawano F., Nishimura H. et al. (1983) A case of K cell deficiency with diabetes mellitus and Graves’ disease. J. Clin. Lab. Immunol., 12(4): 221–224.
McClain K., Gehrz R., Grierson H. et al. (1988) Virus-associated histiocytic proliferations in children. Frequent association with Epstein-Barr virus and congenital or acquired immunodeficiencies. Am. J. Pediatr. Hematol. Oncol., 10(3): 196–205.
Meropol N.J., Porter M., Blumenson L.E. et al. (1996) Daily subcutaneous injection of low-dose interleukin 2 expands natural killer cells in vivo without significant toxicity. Clin. Cancer Res., 2(4): 669–677.
Messina M.F., Cannavo S.P., Aversa S., De Luca F. (2011) Transient natural killer deficiency in a boy with herpes simplex virus-associated recurrent erythema multiforme. Scand. J. Infect. Dis., 43(6–7): 550–552.
Mori T., Yabuhara A., Nakayama J. et al. (1995) Frequently relapsing minimal change nephrotic syndrome with natural killer cell deficiency prior to the overt relapse of Hodgkin’s disease. Pediatr. Nephrol., 9(5): 619–620.
Muhanna N., Abu Tair L., Doron S. et al. (2011) Amelioration of hepatic fibrosis by NK cell activation. Gut, 60(1): 90–98.
Nagao F., Yabe T., Xu M., Okumura K. (1995) Phenotypical and functional analyses of natural killer cells from low NK activity individuals among healthy and patient populations. Nat. Immun., 14(5–6): 225–233.
Nair M.P., Kronfol Z.A., Schwartz S.A. (1990) Effects of alcohol and nicotine on cytotoxic functions of human lymphocytes. Clin. Immunol. Immunopathol., 54(3): 395–409.
Nair M.P., Schwartz S.A. (1984) Immunomodulatory effects of corticosteroids on natural killer and antibody-dependent cellular cytotoxic activities of human lymphocytes. J. Immunol., 132(6): 2876–2882.
Nandakumar S., Woolard S.N., Yuan D. et al. (2008) Natural killer cells as novel helpers in anti-herpes simplex virus immune response. J. Virol., 82(21): 10820–10831.
Nave H., Beutel G., Kielstein J.T. (2011) Obesity-related immunodeficiency in patients with pandemic influenza H1N1. Lancet Infect. Dis., 11(1): 14–15.
Ng S., Fanta C., Okam M., Bhatt A.S. (2011) NK-cell and B-cell deficiency with a thymic mass. N. Engl. J. Med., 364(6): 586–588.
Notarangelo L.D., Mazzolari E. (2006) Natural killer cell deficiencies and severe varicella infection. J. Pediatr., 148(4): 563–564.
Nowak I., Magott-Procelewska M., Kowal A. et al. (2012) Killer immunoglobulin-like receptor (KIR) and HLA genotypes affect the outcome of allogeneic kidney transplantation. PLoS One., 7(9): e44718.
Oosterling S.J., van der Bij G.J., Mels A.K. et al. (2006) Perioperative IFN-alpha to avoid surgically induced immune suppression in colorectal cancer patients. Histol. Histopathol., 21(7): 753–760.
Orange J.S. (2002) Human natural killer cell deficiencies and susceptibility to infection. Microbes Infect., 4(15): 1545–1558.
Orange J.S. (2006) Human natural killer cell deficiencies. Curr. Opin. Allergy Clin. Immunol., 6(6): 399–409.
Orange J.S. (2012) Unraveling human natural killer cell deficiency. J. Clin. Invest., 122(3): 798–801.
Orange J.S. (2013) NK cell deficiency syndromes: Treatment. UpToDate: http://www.uptodate.com/contents/nk-cell-deficiency-syndromes-treatment.
Ostenstad B., Giliani S., Mellbye O.J. et al. (1997) A boy with X-linked hyper-IgM syndrome and natural killer cell deficiency. Clin. Exp. Immunol., 107(2): 230–234.
Ovesen T., Kragelund J.R., Jensen J.M. et al. (2011) Immunodeficiencies in children with chronic post tympanic otorrhoea. Dan. Med. Bull., 58(7): A4282.
Pahwa S., Kirkpatrick D., Ching C. et al. (1983) Persistent cytomegalovirus infection: association with profound immunodeficiency and treatment with interferon. Clin. Immunol. Immunopathol., 28(1): 77–89.
Park Y.W., Kee S.J., Cho Y.N. et al. (2009) Impaired differentiation and cytotoxicity of natural killer cells in systemic lupus erythematosus. Arthritis Rheum., 60(6): 1753–1763.
Platsoucas C.D., Fernandes G., Gupta S.L. et al. (1980) Defective spontaneous and antibody-dependent cytotoxicity mediated by E-rosette-positive and E-rosette-negative cells in untreated patients with chronic lymphocytic leukemia: augmentation by in vitro treatment with interferon. J. Immunol., 125(3): 1216–1223.
Plebani A., Ciravegna B., Ponte M. et al. (2000) Interleukin-2 mediated restoration of natural killer cell function in a patient with Griscelli syndrome. Eur. J. Pediatr., 159(9): 713–714.
Pogorel’skaia E.P., Skriabina E.G., Atmatova G.S. et al. (1985) Quantitative and functional evaluation of immunoregulating cells and natural killers in patients with aplastic anemia. Ter. Arkh., 57(7): 71–74.
Pollock R.E., Lotzova E., Stanford S.D. (1992) Surgical stress impairs natural killer cell programming of tumor for lysis in patients with sarcomas and other solid tumors. Cancer, 70(8): 2192–2202.
Portales P., Reynes J., Pinet V. et al. (2003) Interferon-alpha restores HIV-induced alteration of natural killer cell perforin expression in vivo. AIDS, 17(4): 495–504.
Portaro J.K., Zighelboim J., Fahey J.L. (1978) Hereditary deficiency of K cells in a normal subject. Clin. Immunol. Immunopathol., 11(4): 458–469.
Qiang Q., Zhengde X., Chunyan L. et al. (2012) Killer cell immunoglobulin-like receptor gene polymorphisms predispose susceptibility to Epstein-Barr virus associated hemophagocytic lymphohistiocytosis in Chinese children. Microbiol. Immunol., 56(6): 378–384.
Quesada J.M., Serrano I., Borrego F. et al. (1995) Calcitriol effect on natural killer cells from hemodialyzed and normal subjects. Calcif. Tissue Int., 56(2): 113–117.
Rai N., Thakur N. (2013) Congenital CMV with LAD Type 1 and NK Cell deficiency. J. Pediatr. Hematol. Oncol., Jan. 17 [Epub ahead of print].
Ravaglia G., Forti P., Maioli F. et al. (2000) Effect of micronutrient status on natural killer cell immune function in healthy free-living subjects aged >/=90 y. Am. J. Clin. Nutr., 71(2): 590–598.
Rook A.H., Masur H., Lane H.C. et al. (1983) Interleukin-2 enhances the depressed natural killer and cytomegalovirus-specific cytotoxic activities of lymphocytes from patients with the acquired immune deficiency syndrome. J. Clin. Invest., 72(1): 398–403.
Ruco L.P., Procopio A., Maccallini V. et al. (1983) Severe deficiency of natural killer activity in the peripheral blood of patients with hairy cell leukemia. Blood, 61(6): 1132–1137.
Rukavina D., Laskarin G., Rubesa G. et al. (1998) Age-related decline of perforin expression in human cytotoxic T lymphocytes and natural killer cells. Blood., 92(7): 2410–2420.
Samileh N., Ahmad S., Farzaneh A. et al. (2006) Immunity status in children with Bacille Calmette-Guerin adenitis. A prospective study in Tehran, Iran. Saudi Med. J., 27(11): 1719–1724.
Santos P.C., Falcao R.P. (1990) Decreased lymphocyte subsets and K-cell activity in iron deficiency anemia. Acta Haematol., 84(3): 118–121.
Schleinitz N., Vely F., Harle J.N., Vivier E. (2010) Natural killer cells in human autoimmune diseases. Immunology., 131(4): 451–458.
Schmitt H., Schnitzler N., Riehl J. et al. (1999) Successful treatment of pulmonary Mycobacterium xenopi infection in a natural killer cell-deficient patient with clarithromycin, rifabutin, and sparfloxacin. Clin. Infect. Dis., 29(1): 120–124.
Schneider E.M., Lorenz I., Walther P., Janka-Schaub G.E. (2003) Natural killer deficiency: a minor or major factor in the manifestation of hemophagocytic lymphohistiocytosis? J. Pediatr. Hematol. Oncol., 25(9): 680–683.
Schofer O., Buettner H.J., Kraemer M. et al. (1989) Engraftment of T-depleted major histocompatibility complex-mismatched bone marrow in T-deficient versus natural killer-deficient recipients. Scand. J. Immunol., 29(5): 627–629.
See D.M., Tilles J.G. (1996) Аlpha-interferon treatment of patients with chronic fatigue syndrome. Immunol. Invest., 25(1–2): 153–164.
Shaw R.K., Issekutz A.C., Fraser R. et al. (2012) Bilateral adrenal EBV-associated smooth muscle tumors in a child with a natural killer cell deficiency. Blood, 119(17): 4009–4012.
Shi Y., Lutz C.T. (2002) Interferon-gamma control of EBV-transformed B cells: a role for CD8+ T cells that poorly kill EBV-infected cells. Viral. Immunol., 15(1): 213–225.
Shiozawa K., Shiozawa S. (2006) Interferon alpha (IFNalpha) treatment for Sjogren’s syndrome. Nihon. Rinsho, 64(7): 1345–1353.
Sirianni M.C., Bonomo R., Scarpati B. et al. (1986) Immunological responses of patients with recurrent herpes genitalis. Diagn. Immunol., 4(6): 294–298.
Smith B.R., Rosenthal D.S., Ault K.A. (1985) Natural killer lymphocytes in hairy cell leukemia: presence of phenotypically identifiable cells with defective functional activity. Exp. Hematol., 13(3): 189–193.
Specter S., Rivenbark M., Newton C. et al. (1989) Prevention and reversal of delta-9-tetrahydrocannabinol induced depression of natural killer cell activity by interleukin-2. Int. J. Immunopharmacol., 11(1): 63–69.
Sternfeld T., Nigg A., Belohradsky B.H., Bogner J.R. (2010) Treatment of relapsing Mycobacterium avium infection with interferon-gamma and interleukin-2 in an HIV-negative patient with low CD4 syndrome. Int. J. Infect. Dis., 14(3): e198–201.
Strowig T., Brilot F., Arrey F. et al. (2008) Tonsilar NK Cells Restrict B Cell Transformation by the Epstein-Barr Virus via IFN-?. PLoS Pathog., 4(2): e27.
Sun J.C., Beilke J.N., Lanier L.L. (2009) Adaptive immune features of natural killer cells. Nature, 457(7229): 557–561.
Tapazoglou E., Prasad A.S., Hill G. et al. (1985) Decreased natural killer cell activity in patients with zinc deficiency with sickle cell disease. J. Lab. Clin. Med., 105(1): 19–22.
Thyss A., Caldani C., Bourcier C. et al. (1985) Deficiency of NK activity evolving in a parallel manner with immune granulopenia. Nouv. Rev. Fr. Hematol., 27(6): 383–384.
Toren A., Nagler A., Rozenfeld-Granot G. et al. (2000) Amplification of immunological functions by subcutaneous injection of intermediate-high dose interleukin-2 for 2 years after autologous stem cell transplantation in children with stage IV neuroblastoma. Transplantation, 70(7): 1100–1104.
Toth J., Starsia Z., Buc M., Stefanovic J. (1989) Family study of natural killer cell activity in C1q-deficient patients with systemic lupus erythematosus-like syndrome: association between impaired natural killer cell function and C1qdeficiency. Immunobiology, 180(1): 47–54.
Vaquer S., de la Hera A., Jorda J. et al. (1987) Diminished natural killer activity in pregnancy: modulation by interleukin 2 and interferon gamma. Scand. J. Immunol., 26(6): 691–698.
Villanueva J., Lee S., Giannini E.H. et al. (2005) Natural killer cell dysfunction is a distinguishing feature of systemic onset juvenile rheumatoid arthritis and macrophage activation syndrome. Arthritis Res. Ther., 7(1): R30–37.
Vinh D.C., Patel S.Y., Uzel G. et al. (2010) Autosomal dominant and sporadic monocytopenia with susceptibility to mycobacteria, fungi, papillomaviruses, and myelodysplasia. Blood, 115(8): 1519.
Virelizier J.L. (1984) Deficiencies of interferon production in children. Presse Med., 13(8): 495–498.
Virelizier J.L., Griscelli C. (1981) Selective defect on interferon secretion associated with impaired natural killing activity. Arch. Fr. Pediatr., 38(2): 77–81.
Vossen M.T., Biezeveld M.H., de Jong M.D. et al. (2005) Absence of circulating natural killer and primed CD8+cells in life-threatening varicella. J. Infect. Dis., 191: 198–206.
Wendland T., Herren S., Yawalkar N. et al. (2000) Strong alpha beta and gamma delta TCR response in a patient with disseminated Mycobacterium avium infection and lack of NK cells and monocytopenia. Immunol. Lett., 72(2): 75–82.
Whiteside T.L., Bryant J., Day R., Herberman R.B. (1990) Natural killer cytotoxicity in the diagnosis of immune dysfunction: criteria for a reproducible assay. J. Clin. Lab. Anal., 4(2): 102–114.
Witt M.D., Torno M.S., Sun N., Stein T. (2002) Herpes simplex virus lymphadenitis: case report and review of the literature. Clin. Infect. Dis., 34(1): 1–6.
Witte T., Bode F.M., Hammer M. et al. (1996) Autoantibody against a 58 kD molecule in a patient with neutropenia and NK cell deficiency. Br. J. Haematol., 92(3): 565–570.
Wood S.M., Ljunggren H.G., Bryceson Y.T. (2011) Insights into NK cell biology from human genetics and disease associations. Cell. Mol. Life Sci., 68(21): 3479–3493.
Yang C.A., Raftery M.J., Hamann L. et al. (2012) Association of TLR3-hyporesponsiveness and functional TLR3 L412F polymorphism with recurrent herpes labialis. Hum. Immunol., 73(8): 844–851.
Yang C.M., Yang Y.H., Lin Y.T. et al. (2002) Natural killer cell deficiency associated with Hodgkin’s lymphoma: a case report. J. Formos. Med. Assoc., 101(1): 73–75.
Yeh S., Li Z., Sen H.N. et al. (2010) Scleritis and multiple systemic autoimmune manifestations in chronic natural killer cell lymphocytosis associated with elevated TCRalpha/beta+CD3+CD4–CD8– double-negative T cells. Br. J. Ophthalmol., 94(6): 748–752.
Yoffe B., Pollack S., Ben-Porath E. et al. (1986) Natural killer cell activity in post-necrotic cirrhotic patients as related to hepatitis-B virus infection and plasma zinc levels. Immunol. Lett., 14(1): 15–19.
Zeng W., Maciejewski J.P., Chen G. et al. (2002) Selective reduction of natural killer T cells in the bone marrow of aplastic anaemia. Br. J. Haematol., 119(3): 803–809.
Ziegler H.W., Kay N.E., Zarling J.M. (1981) Deficiency of natural killer cell activity in patients with chronic lymphocytic leukemia. Int. J. Cancer, 27(3): 321–327.
Ziegler-Heitbrock H.W., Rumpold H., Kraft D. et al. (1985) Patients with a deficiency of natural killer cell activity lack the VEP13-positive lymphocyte subpopulation. Blood, 65(1): 65–70.
Zorrilla E.P., Luborsky L., McKay J.R. et al. (2001) The relationship of depression and stressors to immunological assays: a meta-analytic review. Brain. Behav. Immun; 15(3): 199–226.
Дефицит естественных киллеров: гетерогенность, клиника, диагностика, лечение, клинические примеры
Д.В. Мальцев, Я.Я. Недопако
Резюме. Дефицит естественных киллеров — это обобщающий термин, который используется для обозначения группы гетерогенных по происхождению болезней иммунной системы, при которых отмечается уменьшенное количество и/или нарушенная функциональная активность субпопуляции лимфоцитов с фенотипом CD3–СD16+CD56+, однако имеют место нормальные другие показатели иммунного статуса. Можно выделить первичный и вторичный дефицит естественных киллеров. На данный момент описано более 10?первичных форм этого иммунодефицита. В клинике преобладают вирусные инфекции, аутоиммунные и неопластические осложнения, а также некоторые дополнительные симптомы. Для диагностики используют проточную цитометрию, цитологический анализ и ряд функциональных тестов. В качестве базисной терапии применяют препараты интерферонов и рекомбинантного интерлейкина-2. Приведено два клинических случая из собственной практики. У одной пациентки с изолированным дефицитом естественных киллеров развились васкулопатия, обусловленная вирусом ветряной оспы/опоясывающего герпеса, и лейкоэнцефалит, ассоциированный с вирусом герпеса человека 6-го типа. У другой больной сочетался дефицита С3-компонента комплемента и естественных киллеров, что привело к хронической микст-инфекции, вызванной бета-гемолитическим стрептококком и вирусом Эпштейна — Барр, с соответствующими аутоиммунными осложнениями.
.